Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Происхождение и функции клеток иммунной системы
Лимфоциты, как и другие клетки крови, ведут свое происхождение из единой стволовой гемопоэтической клетки. Лимфопоэз имеет черты сходства с общим гемопоэзом: зависит от контактов клеток-предшественниц со стромальными клетками и от влияния цитокинов. Вместе с тем лимфопоэзу присущи некоторые особенности: возникновение в процессе пролиферации клеток-предшественниц широкого разнообразия Т- и В-лимфоцитов с разными антигенраспознающими рецепторами; гибель путем апоптоза значительной части пролиферирующих клеток-предшественниц; избирательное накопление в результате пролиферации клеток — потомков одной клетки, первоначально активированной при контакте с соответствующим антигеном, «клона» клеток, все клетки которого несут одинаковые по специфичности антигенраспознающие рецепторы. Лимфоциты — это единственные клетки организма, способные специфически распознавать собственные и чужеродные антигены и отвечать активацией на контакт с конкретным антигеном. При весьма сходной морфологии малые лимфоциты делятся на две популяции, имеющие различные функции и продуцирующие разные белки (табл. 8.1). Одна из популяций получила название В-лимфоциты, от названия органа «бурса Фабрициуса», где было впервые обнаружено созревание этих клеток у птиц. У человека В-лимфоциты созревают в костном мозге. В- лимфоциты распознают антигены специфическими рецепторами иммуноглобулиновой природы, которые по мере созревания В-лимфоцитов появляются на их мембранах. Взаимодействие антигена с такими рецепторами является сигналом активации В-лимфоцитов и их дифференцировки в плаз- Таблица 8.1. Классификация лимфоцитов человека
матические клетки, продуцирующие и секретирующие специфические для данного антигена антитела — иммуноглобулины.
Другая популяция получила название Т-лимфоциты в связи с дифференцировкой их предшественников в тимусе. Эта популяция включает разные субпопуляции, среди которых наиболее отчетливо различаются Т-хелперы (CD4+) и цитотоксические Т-лимфоциты (CD8+). В отличие от В-лим- фоцитов, способных распознавать и связывать белковые, полисахаридные и липопротеидные растворимые антигены, Т-лимфоциты могут распознавать только короткие пептидные фрагменты белковых антигенов на поверхности вспомогательных антигенпредставляющих клеток (дендритных, макрофагов или В-лимфоцитов) в комплексах с собственными молекулами главного комплекса гистосовместимости. Связывание антигена с антиген- распознающим рецептором на клеточной мембране индуцирует активацию Т-лимфоцита, которая проявляется продукцией и секрецией цитокинов, усиливающих процессы пролиферации и дифференцировки самих Т-лимфоцитов, В-лимфоцитов и макрофагов. Т-лимфоциты выполняют важнейшую функцию специфического распознавания и связывания антигена, что ведет к их активации, пролиферации и дифференцировке разных субпопуляций Т-лимфоцитов, участвующих далее во всех формах иммунного ответа. Главной функцией В-лимфоцитов также является специфическое распознавание антигена, которое ведет к их активации, пролиферации и дифференцировке в плазматические клетки — продуценты специфических антител — иммуноглобулинов, т. е. к гуморальному иммунному ответу. Чаще всего В-лимфоциты для развития гуморального иммунного ответа нуждаются в помощи Т-лимфоцитов в виде продукции активирующих цитокинов. Имеется еще один тип лимфоцитов — большие гранулярные лимфоциты, отличающиеся от меньших по величине Т- и В-лимфоцитов не только особенностями структуры, но и отсутствием антигенраспознающего рецептора. Эти клетки получили название «естественные киллеры»: они способны убивать инфицированные разными вирусами клетки-мишени или опухолевые клетки (см. табл. 8.1).
Все процессы транспортировки лимфоцитов из первичных во вторичные органы иммунной системы, из вторичных органов иммунной системы в ткани зависят от взаимодействия особого типа цитокинов — хемокинов с соответствующими рецепторами на поверхности клеток. Кроме хемокинов миграция лимфоцитов зависит от присутствия на их поверхности адгезионных молекул, которым соответствуют молекулы-лиганды на поверхности эндотелиальных клеток сосудов, на поверхности макрофагов и дендритных клеток, на элементах внеклеточного матрикса. 8.1.1. Т-лимфоциты 8.1.1.1. Характеристики Т-лимфоцитов Решающее событие в развитии Т-лимфоцитов — формирование антигенраспознающего Т-клеточного рецептора — происходит только в тимусе. Для обеспечения возможности распознавания любого антигена нужны миллионы различных по специфичности антигенраспознающих рецепторов. Формирование огромного разнообразия антигенраспознающих рецепторов возможно благодаря перестройке генов в процессе пролиферации и дифференцировки клеток-предшественниц. Таблица 8.2. Типы молекул на поверхности Т-лимфоцитов
По мере созревания Т-лимфоцитов на их поверхности появляются ан- тигенраспознающие рецепторы и другие молекулы, опосредующие их взаимодействие с антигенпредставляющими клетками. Так, в распознавании собственных молекул главного комплекса гистосовместимости (см. раздел 8.3.2) наряду с Т-клеточным рецептором участвуют молекулы CD4 или CD8. Межклеточные контакты обеспечиваются наборами поверхностных адгезионных молекул, каждой из которых соответствует молекула — лиганд на поверхности другой клетки. Как правило, взаимодействие Т- лимфоцита с антигенпредставляющей клеткой не ограничивается распознаванием антигенного комплекса Т-клеточным рецептором, а сопровождается связыванием других попарно комплементарных поверхностных «костимулирующих» молекул. Сочетание поверхностных молекул лимфоцитов, которые принято обозначать порядковыми номерами «кластеров дифференцировки» (clusters of differentiation — CD), обозначается как «поверхностный фенотип клетки», а отдельные поверхностные молекулы называют «маркерами», так как они служат метками конкретных субпопуляций и стадий дифференцировки Т- лимфоцитов. Так, например, на поздних этапах дифференцировки одни Т- лимфоциты утрачивают молекулу CD8 и сохраняют только CD4, а другие утрачивают CD4, а сохраняют CD8. Поэтому среди зрелых Т-лимфоцитов различают CD4+ (Т-хелперы) и CD8+ (цитотоксические Т-лимфоциты). Среди циркулирующих в крови Т-лимфоцитов клеток с маркером CD4 примерно в два раза больше, чем клеток с маркером CD8.
Зрелые Т-лимфоциты несут на поверхности рецепторы для разных цитокинов и рецепторы для иммуноглобулинов (табл. 8.2). При распознавании Т-клеточным рецептором антигена Т-лимфоциты получают сигналы активации, пролиферации и дифференцировки в направлении клеток-эффекторов, т. е. клеток, способных непосредственно участвовать в защитных или повреждающих эффектах. Для этого на их поверхности резко возрастает количество адгезионных и костимулирующих молекул, а также рецепторов для цитокинов. Активированные Т-лимфоциты начинают продуцировать и секретировать цитокины, активирующие макрофаги, другие Т-лимфоциты и В-лимфоциты. После завершения инфекции, сопряженной с усиленной продукцией, дифференцировкой и активацией Т-эффекторов соответствующего клона, в течение нескольких дней 90 % эффекторных клеток погибают, поскольку не получают дополнительных сигналов активации. В организме остаются долгоживущие клетки памяти, несущие соответствующие по специфичности рецепторы и способные ответить пролиферацией и активацией на повторную встречу с тем же антигеном. 8.1.1.2. Субпопуляции Т-лимфоцитов Основная функция CD4+ Т-лимфоцитов — продуцировать цитокины (см. раздел 8.4.2), активирующие функции других клеток, в связи с этим данные клетки называют помощниками (helper) и обозначают соответственно Th. Основной функцией СЕ)8+Т-лимфоцитов является цитотоксичность, т. е. способность убивать клетки-мишени, инфицированные соответствующим по специфичности вирусом, поэтому их называют цитотоксическими Т-лимфоцитами и обозначают CTL (табл. 8.3). Таблица 8.3. Характеристики трех типов эффекторных Т-лимфоцитов
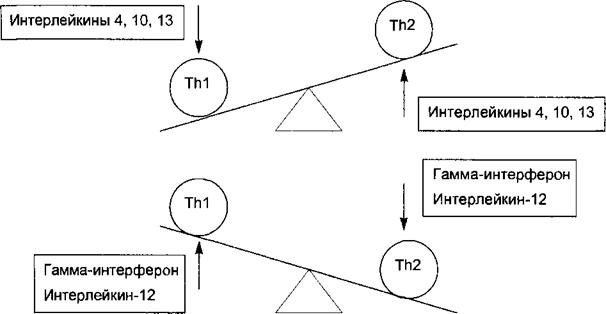
С04+Т-лимфоциты после распознавания антигена начинают секретировать интерлейкины (ИЛ-2, ИЛ-4), гамма-интерферон и дифференцируются в двух направлениях. Одни — Thl — приобретают способность активировать макрофаги и участвовать в клеточном иммунном ответе. Другое направление дифференцировки ведет к формированию Th2, способных акти-
Рис. 8.2. Цитокиновый баланс, от которого зависит преобладание клеточного или гуморального иммунного ответа. Превалирование Th2 приводит к усилению гуморального ответа и подавлению ТЫ. Превалирование ТЫ приводит к усилению клеточного иммунного ответа и подавлению Th2. Эти альтернативные эффекты опосредованы разными группами цитокинов.
вировать В-лимфоциты к продукции соответствующих по специфичности антител, т. е. участвуют в гуморальном иммунном ответе. Каждая из двух субпопуляций Th продуцирует аутокринно и паракринно действующие цитокины, способные помешать дифференцировке и активации альтернативной субпопуляции (рис. 8.2). 8.1.1.3. Функции Т-лимфоцитов При развитии специфического иммунного ответа Т-лимфоциты в лимфатических узлах, селезенке и мукозноассоцииросванных лимфоидных тканях выполняют секреторные и эффекторные функции. Активированные Т-лимфоциты продуцируют и секретируют молекулы цитокинов. Цитокины связываются со специфическими рецепторами на поверхности клеток-мишеней. Соединение цитокина с его рецептором порождает сигнал активации, который передается соответствующими факторами трансдукции к ядру клетки-мишени, где начинают функционировать определенные гены, контролирующие функции клеток. Таким образом, Т- лимфоциты своими продуктами цитокинами контролируют процессы пролиферации, дифференцировки и функциональную активность макрофагов, дендритных клеток, других Т-лимфоцитов, В-лимфоцитов. Некоторые цитокины непосредственно участвуют в защите организма: • интерфероны обладают противовирусным действием, • туморнекротизирующий фактор (ТНФ) оказывает цитотоксическое действие на некоторые опухолевые клетки. Активированные CD8+ Т-лимфоциты выполняют эффекторную функцию цитотоксических Т-лимфоцитов (CTL): распознают своими рецепторами и убивают клетки-мишени, несущие на своей поверхности соответствующий по специфичности антигенный пептид. При непосредственном контакте CTL с клеткой-мишенью содержимое гранул CTL (цитотоксины: перфорины и грензимы) проникают в клетку-мишень и вызывают ее гибель. 8. L2. В-лимфоциты 8.1.2.1. Характеристики В-лимфоцитов Предшественники В-лимфоцитов из эмбриональной печени человека на 8—9-й неделе развития плода перемещаются в костный мозг, где они созревают под влиянием интерлейкина 7, который секретируют стромальные клетки. В ранний период созревания в костном мозге формируется и фиксируется на поверхностной мембране В-лимфоцита иммуноглобулиновый анти- генраспознающий рецептор. Незрелый В-лимфоцит уже несет мембранные В- клеточные рецепторы (IgM) определенной специфичности, т. е. способные распознать и связать конкретный антиген. При встрече незрелых В-лимфоцитов с аутоантигенами связывание последних с мембранными IgM ведет к остановке дифференцировки этих В-лимфоцитов, с последующей их гибелью путем апоптоза. На этой стадии погибают до 90 % незрелых В- клеток. Часть незрелых В-клеток после контакта с растворимым аутоантигеном не погибают, а переходят в состояние анергии по отношению к данному антигену. Этим объясняется врожденное состояние иммунологической толерантности (неспособности ответить) на собственные аутоантигены.
Выжившие в этом процессе отбора В-клетки по мере созревания покидают костный мозг, переходя в кровяное русло через венозные синусы, и поступают в периферические органы иммунной системы, где превращаются в зрелые В-лимфоциты. В кровяном русле В-лимфоциты включаются в рециркуляцию и заселяют В-зоны периферических органов и тканей иммунной системы. Около 2 • 107 В-лимфоцитов покидают костный мозг ежедневно, что составляет 5—10 % общей популяции В-клеток периферической крови. Этого достаточно, чтобы полностью обновить популяцию периферических В-лимфоцитов за 4—5 дней. Среди сохранившихся и вступивших в рециркуляцию относительно долгоживущих В-лимфоцитов (с полупериодом жизни 1—2 мес) значительную часть составляют клетки памяти, способные быстро ответить пролиферацией, активацией, продукцией и секрецией специфических антител (иммуноглобулинов) на повторную встречу с тем же антигеном. 8.1.2.2. Функции В-лимфоцитов В-лимфоциты с помощью своих иммуноглобулиновых рецепторов могут распознавать не только пептиды, но и более крупные молекулы белков, нуклеиновых кислот, углеводов, липидов, полисахаридные и липопротеидные растворимые антигены. Зрелые В-лимфоциты способны самостоятельно распознавать растворимые антигены, в отличие от Т-лимфоцитов они не нуждаются в презентации чужеродных антигенов. Более того, В-лимфоциты сами могут выполнять функцию антигенпредставляющих клеток при взаимодействии с Th2. Для этого В-лимфоциты имеют на своей поверхности адгезионные и костимулирующие молекулы, рецепторы для иммуноглобулинов, компонентов системы комплемента и для цитокинов (табл. 8.4). Распознавание антигена В-клеточным рецептором служит одним из сигналов активации В-лимфоцитов, которая проявляется их пролиферацией и дифференцировкой в продуцирующие иммуноглобулины плазматические клетки. Дополнительные сигналы активации В-клетки получают при взаимодействии с Th2, которые продуцируют и секретируют соответствующие цитокины: интерлейкины 4, 5, 6, 10. Синтез и секреция антител (иммуноглобулинов) В-лимфоцитами является завершающей стадией специфического гуморального иммунного ответа на конкретный антиген. В динамике иммунного ответа происходит переключение синтеза антител, который начинается с IgM, на другие изотипы: Таблица 8.4. Типы молекул на поверхности В-лимфоцитов
IgG, IgA или IgE. Переключение индуцируется разными цитокинами и взаимодействием костимулирующих молекул (CD40—CD40L), присутствующих на поверхности В-лимфоцитов и Th2. 8.1.3. Антигенпредставляющие клетки Способностью представлять (презентировать) антигенные пептиды Т-лимфоцитам обладают антигенпредставляющие клетки: дендритные клетки, макрофаги и В-лимфоциты. Презентации антигенных пептидов предшествуют стадии: 1) захвата поступившего в организм антигена, 2) его переработки (дезинтеграции), 3) формирования комплексов накопившихся антигенных пептидов с собственными молекулами главного комплекса гистосовместимости, постоянно синтезирующиеся в этих клетках (см. раздел 8.3.2), 4) транспортировки образовавшихся комплексов на мембрану антигенпрезентирующей клетки, 5) доставки во вторичные лимфоидные органы, где и происходит встреча с Т-лимфоцитами и распознавание образовавшегося комплекса Т-клеточным рецептором. Дендритные клетки, как и макрофаги и лимфоциты, имеют гемопоэтическое происхождение. Дендритные клетки локализованы в эпителии кишечника, урогенитального тракта, воздухоносных путей, легких, в эпидермисе кожи (клетки Лангерганса) и интерстициальных пространствах.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-09; просмотров: 76; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.43.122 (0.027 с.) |