Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Каковы особенности структурной организации и функции нуклеиновых кислот. В чем отличие днк и рнк, их свойства. В чем особенности строения атф.
Нуклеиновые кислоты - это линейные неразветвленные гетерополимеры, мономерами которых являются нуклеотиды,связанные фосфоэфирными связями. Существует два типа нуклеиновых кислот: ДНК (дезоксорибонуклеиновая кислота) и РНК (рибонуклеиновая кислота). Нуклеиновые кислоты обеспечивают хранение, воспроизведение и реализацию генетической (наследственной) информации. Эта информация отражена (закодирована) в виде нуклеотидных последовательностей. В частности, последовательность нуклеотидов отражает первичную структуру белков. Соответствие между аминокислотами и кодирующими их нуклеотидными последовательностями называется генетическим кодом. Единицей генетического кода ДНК и РНК является триплет-последовательность из трех нуклеотидов. Нуклеиновые кислоты образуют разнообразные соединения с белками - нуклеопротеины или нуклеопротеиды ДНК – это нуклеиновая кислота, мономерами которой являются дезоксирибонуклеотиды.
В клетках эукариот (например, животных или растений) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одно- или двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих вирусов.
С химической точки зрения, ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков, нуклеотидов.
Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы.
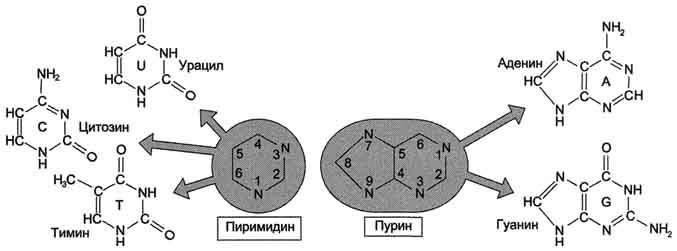
Пуриновые и пиримидиновые основания.
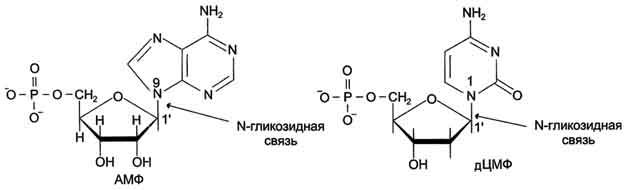
Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы Каждый нуклеотид состоит из остатка фосфорной кислоты присоединённого по 5'-положению к сахару дезоксирибозе, к которому также через гликозидную связь (C—N) по 1'-положению присоединено одно из четырёх азотистых оснований..
Пуриновый и пиримидиновый нуклеотиды.
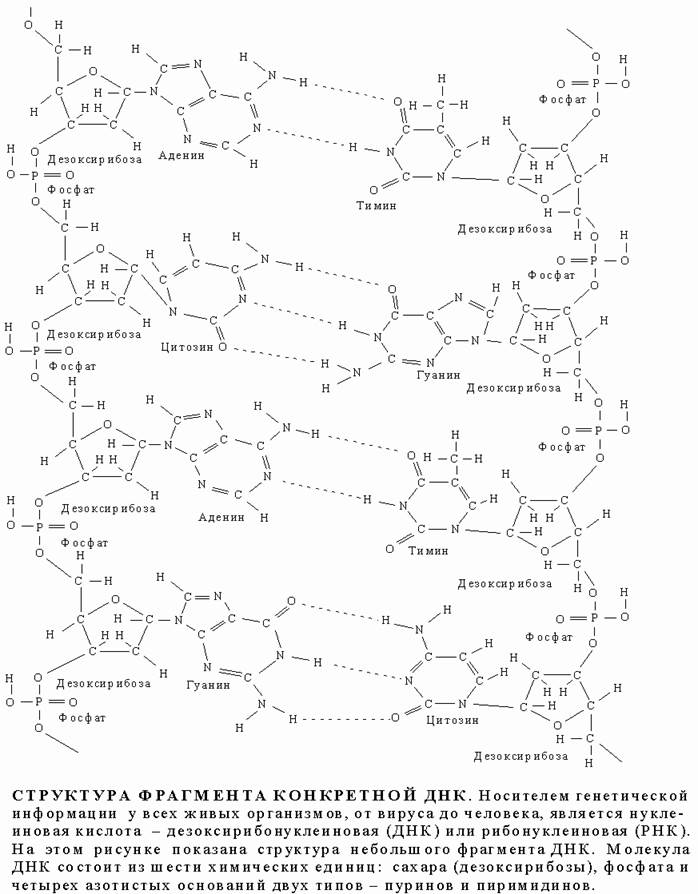
Фрагмент цепи ДНК.
В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула спирализована. В целом структура молекулы ДНК получила название «двойной спирали».
В 1953 г. Дж. Уотсоном и Ф. Криком была предложена модель пространственной структуры ДНК. Согласно этой модели, молекула ДНК имеет форму спирали, образованную двумя полинуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Двойная спираль правозакрученная, полинуклеотидньхе цепи в ней антипараллельны, т.е. если одна из них ориентирована в направлении 3'→5', то вторая - в направлении 5'→3'. Поэтому на каждом из концов
В ДНК встречается четыре вида азотистых оснований (аденин, гуанин, тимин и цитозин). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин соединяется только с тимином, гуанин — только с цитозином.
ДНК является первичным носителем наследственной информации. Это означает, что вся информация о структуре, функционировании и развитии отдельных клеток и целостного организма записана в виде нуклеотидных последовательностей ДНК. Обычно молекула ДНК рассматривается как двойная правозакрученная спираль (толщиной ~ 1,8 нм), которая состоит из двух цепей (нитей), связанных между собой водородными связями. Каждая цепь представлена чередующимися остатками дезоксирибозы и фосфорной кислоты, причем, к дезоксирибозе ковалентно присоединяется азотистое основание. При этом азотистые основания двух нитей ДНК направлены друг к другу и за счет образования водородных связей образуют комплементарные пары: А=Т (две водородных связи) и Г=_Ц (три водородных связи). Поэтому нуклеотидные последовательности этих цепей однозначно соответствуют друг другу. Остов каждой из цепей состоит из чередующихся фосфатов и сахаров Фосфатные группы формируют фосфодиэфирные связи между третьим и пятым атомами углерода соседних молекул дезоксирибозы в результате взаимодействия между 3'-гидроксильной (3'—ОН) группой одной молекулы дезоксирибозы и 5'-фосфатной группой (5'—РО3) другой. Асимметричные концы цепи ДНК называются 3' (три прим) и 5' (пять прим). Полярность цепи играет важную роль при синтезе ДНК (удлинение цепи возможно только путём присоединения новых нуклеотидов к свободному 3'-концу).
Ширина двойной спирали составляет от 22 до 24 Å, или 2,2 — 2,4 нм, длина каждого нуклеотида 3,3 Å (0,33 нм). Подобно тому, как в винтовой лестнице сбоку можно увидеть ступеньки, на двойной спирали ДНК в промежутках между фосфатным остовом молекулы можно видеть рёбра оснований, кольца которых расположены в плоскости, перпендикулярной по отношению к продольной оси макромолекулы. В двойной спирали различают малую (12 Å) и большую (22 Å) бороздки. Белки, например, факторы транскрипции, которые присоединяются к определённым последовательностям в двухцепочечной ДНК, обычно взаимодействуют с краями оснований в большой бороздке, где те более доступны. Каждое основание на одной из цепей связывается с одним определённым основанием на второй цепи. Такое специфическое связывание называется Пурины комплементарны пиримидинам (то есть, способны к образованию водородных связей с ними): аденин образует связи только с тимином, а цитозин — с гуанином. В двойной спирали цепочки также связаны с помощью гидрофобных связей и стэкинга, которые не зависят от последовательности оснований ДНК. Комплементарность двойной спирали означает, что информация, содержащаяся в одной цепи, содержится и в другой цепи. Обратимость и специфичность взаимодействий между комплементарными парами оснований важна для репликации ДНК и всех остальных функций ДНК в живых организмах. Так как водородные связи нековалентны, они легко разрываются и восстанавливаются. Цепочки двойной спирали могут расходиться как замок-молния под действием ферментов (хеликазы) или при высокой температуре. Разные пары оснований образуют разное количество водородных связей. АТ связаны двумя, ГЦ — тремя водородными связями, поэтому на разрыв ГЦ требуется больше энергии. Процент ГЦ пар и длина молекулы ДНК определяют количество энергии, необходимой для диссоциации цепей: длинные молекулы ДНК с большим содержанием ГЦ более тугоплавки. Длина ДНК измеряется числом нуклеотидных пар. Длина одной молекулы ДНК колеблется от нескольких тысяч до нескольких миллионов нуклеотидных пар. Например, у наиболее простых вирусов длина ДНК составляет примерно 5 тысяч нуклеотидных пар, у наиболее сложных вирусов – свыше 100 тысяч нуклеотидных пар, у бактерий 4 миллиона нуклеотидных пар, у дрожжей – 13,5 миллиона нуклеотидных пар у мушки дрозофилы – 105 миллиона нуклеотидных пар, у человека – 2900 миллиона нуклеотидных пар (размеры ДНК даны для минимального набора хромосом – гаплоидного). При этом длина участка ДНК, соответствующая одной паре нуклеотидов ~ 0,34 нм; тогда длина ДНК в хромосоме бактерии ~ 1,5 мм, а в гаплоидном наборе человека ~ 1 м.
Объёмные модели ДНК в В- и Z-формах (жирной линией обозначен сахарофосфатный остов).
Репликация ДНК Реплика́ция ДНК — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты, идущий во время синтетической (S) фазы жизненного цикла клетки на матрице родительской молекулы ДНК. При этом генетический материал, зашифрованный в ДНК, удваивается и в процессе последующего деления делится между дочерними клетками. Репликацию ДНК осуществляет сложный ферментный комплекс, состоящий из 15-20 различных белков.
Репликация (самоудвоение) ДНК - это один из важнейших биологических процессов, обеспечивающих воспроизведение генетической информации. В результате репликации одной молекулы ДНК образуется две новые молекулы, которые являются точной копией исходной молекулы- матрицы. Каждая новая молекула состоит из двух цепей – одной из родительских и одной из сестринских. Такой механизм репликации ДНК называется полуконсервативным. Реакции, в которых одна молекула гетерополимера служит матрицей (формой) для синтеза другой молекулы гетерополимера с комплементарной структурой, называются реакциями матричного типа. Если в ходе реакции образуются молекулы того же вещества, которое служит матрицей, то реакция называется автокаталитической. Если же в ходе реакции на матрице одного вещества образуются молекулы другого вещества, то такая реакция называется гетерокаталитической. Таким образом, репликация ДНК (то есть синтез ДНК на матрице ДНК) являетсяавтокаталитической реакцией матричного синтеза.
Кроме репликации ДНК к реакциям матричного типа относятся транскрипция ДНК (синтез РНК на матрице ДНК) и трансляция РНК (синтез белков на матрице РНК). Существуют и другие реакции матричного типа, например, синтез РНК на матрице РНК и синтез ДНК на матрице РНК. Два последних типа реакций наблюдаются при заражении клетки определенными вирусами. Синтез ДНК на матрице РНК (обратная транскрипция) широко используется в генной инженерии.
Все матричные процессы состоят из трех этапов:
Репликация ДНК – это сложный процесс, в котором принимает участие несколько десятков ферментов. К важнейшим из них относятся ДНК-полимеразы (несколько типов), праймазы, топоизомеразы, лигазы и другие.
Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая нить, (3) ДНК полимераза (Polα), (4) ДНК лигаза, (5) РНК праймер, (6) ДНК праймаза, (7) фрагмент Оказаки, (8) ДНК полимераза (Polδ), (9) хеликаза, (10) одиночная нить со связанными белками, (11) топоизомераза
Репликация ДНК – процесс матричный. Так как две цепочки молекулы ДНК комплементарны друг другу (т.е. азотистые основания дополняют друг друга), то каждая из них может служить шаблоном или матрицей для построения новых цепей.
Для удвоения ДНК необходимы: материал, из которого будут строиться новые молекулы ДНК; энергия, т.е. молекулы АТФ; ферменты; матрица – основа, по которой строятся новые полинуклеотидные цепочки.
1. Реплицируется не одна, а обе цепи ДНК каждой хромосомы. 2. Обе цепи ДНК реплицируются полностью — от одного конца до другого 3. ДНК-полимераза представляет собой комплекс основных ферментов репликации. Этот комплекс прикрепляется к ДНК и начинает двигаться вдоль нее. ДНК-полимеразы последовательно наращивают одноцепочечную цепь ДНК, шаг за шагом присоединяя к ней следующие звенья в направлении от 5'-концу к 3'-концу, причем выбор очередного дНТФ диктуется матрицей.Другой фермент — ДНК-лигаза, который катализирует образование связей между соседними нуклеотидами, используя для этого энергию фосфатных связей. ДНК-лигазы сшивают дезоксирибонуклеотиды в цепочку 4. Дочерние цепи ДНК начинают формироваться одновременно в сотнях участков обеих родительских цепей. Впоследствии концы отдельных сегментов вновь синтезированной ДНК сшиваются ферментом ДНК-лигазой. Репликация происходит путем непрерывного роста нуклеотида за нуклеотидом обеих новых цепей по мере перемещения репликационной вилки; при этом, поскольку две цепи в спирали ДНК антипараллельны, о матрица считывается ДНК-полимеразой в направлении 3'–5'. Ссинтез ДНК происходит непрерывно только на одной из матричных цепей. На второй матричной цепи ДНК синтезируется сравнительно короткими фрагментами (длиной от 100 до 1000 нуклеотидов, в зависимости от вида), названными по имени обнаружившего их ученого фрагментами Оказаки. Вновь образованная цепь, которая синтезируется непрерывно, называется ведущей, а другая, собираемая из фрагментов Оказаки, – отстающей. Синтез каждого из этих фрагментов начинается с РНК-затравки. Через некоторое время РНК-затравки удаляются, бреши застраиваются ДНК-полимеразой и фрагменты сшиваются в одну непрерывную цепь ДНК специальным ферментом. Это так называемый прерывистый синтез ДНК (другой учебник) Главная проблема при репликации ДНК заключается в том, что в разных цепях одной молекулы остатки фосфорной кислоты направлены в разные стороны, но наращивание цепей может происходить только с того конца, который заканчивается группой ОН. Поэтому в реплицируемом участке, который называется вилкой репликации, процесс репликации протекает на разных цепях по-разному. На одной из цепей, которая называется ведущей, происходит непрерывный синтез ДНК на матрице ДНК. На другой цепи, которая называется запаздывающей, вначале происходит связывание праймера - специфического фрагмента РНК. Праймер служит затравкой для синтеза фрагмента ДНК, который называется фрагментом Оказаки. В дальнейшем праймер удаляется, а фрагменты Оказаки сшиваются между собой в единую нить фермента ДНК–лигазы.)
5. Каждая вновь синтезированная цепь ДНК остается прикрепленной посредством слабых водородных связей к родительской цепи, используемой в качестве матрицы. Впоследствии обе цепи ДНК вместе скручиваются в спираль. 6. Каждая цепь ДНК имеет длину около 6 см и состоит из миллионов витков, поэтому раскрутить две цепи без специального механизма было бы невозможно. Это достигается с помощью ферментов, которые регулярно разрезают каждую спираль по всей длине, поворачивают ее фрагменты так, чтобы они могли расплестись, и затем вновь восстанавливают целостность каждой спирали. Так возникают две новые спирали.
СМ НИЖЕ РИСУНОК Репликация ДНК. А. Вилка репликации. Новая нить ДНК синтезируется только в направлении от 5'- к З'-концу. Каждая из двух нитей ДНК служит матрицей для синтеза новой нити. Так как родительские нити антипараллельны, то непрерывная репликация ДНК происходит в направлении 5' -> 3' только на одной нити, которая называется ведущей (лидирующей). Б. Синтез новой цепи на отстающей нити требует постоянного образования новых затравок для начала репликации и осуществляется небольшими сегментами по 1000—2000 нуклеотидов в каждом (фрагменты Оказаки). Заправки представляют собой короткие последовательности РНК, которые синтезируются при участии РНК-полимеразы (праймазы). Затравки деградируют после завершения синтеза следующего фрагмента Оказаки. Образованные соседние фрагменты ДНК соединяются ДНК-лигазой. В. Показано, как происходит движение репликативной вилки. Топоизоме-раза удаляет супервитки спирали, хеликаза обеспечивает раскручивание двойной спирали, белок SSB обеспечивает стабильность одноцепочечной ДНК
Репликация ДНК сопровождается репарацией - исправлением ошибок, неизбежно возникающих при репликации. Репарация — особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённой при нормальном биосинтезе ДНК в клетке или в результате воздействия физическими или химическими агентами. Осуществляется специальными ферментными системами клетки. Ряд наследственных болезней (напр., пигментная ксеродерма) связан с нарушениями систем репарации.
РНК – это нуклеиновая кислота, мономерами которой являются рибонуклеотиды. Нуклеиновые кислоты (РНК) — нуклеиновые кислоты, полимеры нуклеотидов, в состав которых входят остаток ортофосфорной кислоты, рибоза (в отличие от ДНК, содержащей дезоксирибозу) и азотистые основания — аденин, цитозин, гуанин и урацил (в отличие от ДНК, содержащей вместо урацила тимин). Эти молекулы содержатся в клетках всех живых организмов, а также в некоторых вирусах.
Нуклеотиды РНК состоят из сахара — рибозы, к которой в положении 1' присоединено одно из оснований: аденин, гуанин, цитозин или урацил. Фосфатная группа соединяет рибозы в цепочку, образуя связи с 3' атомом углерода одной рибозы и в 5' положении другой. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК — полианион. РНК транскрибируется как полимер четырёх оснований (аденина (A), гуанина (G), урацила (U) и цитозина (C)), но в «зрелой» РНК есть много модифицированных оснований и сахаров Всего в РНК насчитывается около 100 разных видов модифицированных нуклеозидов.Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом. Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырёх нуклеотидов, в которой есть пара оснований аденин — гуанин Важная структурная особенность РНК, отличающая её от ДНК — наличие гидроксильной группы в 2' положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, наиболее часто наблюдаемой у ДНК У А-формы глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка. Второе последствие наличия 2' гидроксильной группы состоит в том, что конформационно пластичные, то есть не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять Рабочая форма одноцепочечной молекулы РНК, как и у белков, часто обладает третичной структурой. Третичная структура образуется на основе элементов вторичной структуры, образуемой с помощью водородных связей внутри одной молекулы. Различают несколько типов элементов вторичной структуры — стебель-петли, петли и псевдоузлы. В силу большого числа возможных вариантов спаривания оснований предсказание вторичной структуры РНК — гораздо более сложная задача, чем предсказание вторичной структуры белков, но в настоящее время есть эффективные программы.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (мРНК) принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК. Для одноцепочечных РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитической основой рибосом. Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так малые ядерные РНК принимают участие в сплайсинге эукариотических матричных РНК и других процессах. Помимо того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы) у отдельных РНК обнаружена собственная энзиматическая активность, способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами. Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.
Многие типы РНК, например, рРНК и мяРНК в клетке функционируют в виде комплексов с белками, которые ассоцииируют с молекулами РНК после их синтеза или (у эукариот) экспорта из ядра в цитоплазму. Такие РНК-белковые комплексы называются рибонуклеопротеиновыми комплексами или рибонуклеопротеидами. В клетках обнаруживается три основных типа РНК, выполняющих различные функции: 1. Информационная или матричная РНК (иРНК, или мРНК). Составляет 5% клеточной РНК. Служит для передачи генетической информации от ДНК на рибосомы при биосинтезе белка. В эукариотических клетках иРНК (мРНК) стабилизирована с помощью специфических белков. Это делает возможным продолжение биосинтеза белка даже в том случае, если ядро неактивно. 2. Рибосомная или рибосомальная РНК (рРНК). Составляет 85% клеточной РНК. Входит в состав рибосом, определяет форму большой и малой рибосомных субъединиц, обеспечивает контакт рибосомы с другими типами РНК. 3. Транспортная РНК (тРНК). Составляет 10% клеточной РНК. Транспортирует аминокислоты к соответствующему участку иРНК в рибосомах. Каждый тип тРНК транспортирует определенную аминокислоту. 4. Минорные (малые) РНК. Это молекулы РНК, чаще всего с небольшой молекулярной массой, располагающиеся в различных участках клетки (мембране, цитоплазме, органеллах, ядре и т.д.). Их роль до конца не изучена. Доказано, что они могут помогать созреванию рибосомной РНК, участвуют в переносе белков через мембрану клетки, способствуют редупликации молекул ДНК и т.д. 5. Рибозимы. Недавно выявленный вид РНК, принимающие активное участие в ферментативных процессах клетки в качестве фермента (катализатора). 6. Вирусные РНК. Любой вирус может содержать только один вид нуклеиновой кислоты: либо ДНК либо РНК. Соответственно, вирусы, имеющие в своём составе молекулу РНК, получили название РНК-содержащие. При попадании в клетку вируса данного типа может происходить процесс обратной транскрипции (образование новых ДНК на базе РНК), и уже вновь образовавшаяся ДНК вируса встраивается в геном клетки и обеспечивает существование, а также размножение возбудителя. Вторым вариантом сценария является образование комплиментарной РНК на матрице поступившей вирусной РНК. В этом случае, образование новых вирусных белков, жизнедеятельность и размножение вируса происходит без участия дезоксирибонуклеиновой кислоты только на основании генетической информации, записанной на вирусной-РНК.
Кроме того, в клетках имеются и другие типы РНК, выполняющие вспомогательные функции. Все типы РНК образуется в результате реакций матричного синтеза. В большинстве случаев матрицей служит одна из цепей ДНК. Таким образом, синтез РНК на матрице ДНК является гетерокаталитической реакцией матричного типа. Этот процесс называется транскрипцией и контролируется определенными ферментами – РНК–полимеразами (транскриптазами). «Сравнительная характеристика ДНК и РНК»
|
||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-19; просмотров: 750; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.218.234.83 (0.049 с.) |