Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Джерела формуванняя і поповнення генофонду вірусних популяцій
4. Мутації, які мають фенотипові прояви (наприклад, зміни розмірів бляшок, термостабільність). В природних умовах точкові мутації генів гемаглютиніну і нейромінідази обумовлюють антигенний дрейф вірусів грипу (зміни структури поверхневих антигенів) в динаміці епідемічного процесу. Найкраще вивчені умовно-летальні мутанти; у яких один з вірусних білків втрачає здатність функціюнувати або синтезується лише за визначених умов. Прикладом мутантів з дефектом індивідуального гена є холодові мутанти. Вони не здатні репродукуватися в інтервалі температур 37-410С. Їх широко використовують для виготовлення вакцин або активної імунізації. Мутації, які збільшують інфекційний спектр, обумовлюють здатність репродукуватися в клітинах, що є нечутливими до “диких” штамів вірусу. Мутації, що обумовлюють стійкість до антивірусних прапаратів є, характерними для РНК- та ДНК-вмісних вірусів. Термостабільні мутанти здатні реплікуватися при 410С і виключно вірулентні. Бляшкоутворюючі мутанти характеризуються зміною розмірів бляшок (зон лізісу) в моношарі клітинної культури або на бактеріальному газоні. Мутації, що обумовлюють дефіцит ферменту, викликають повну втрату ферменту або модифікацію його структури, Мутації можуть бути летальними або умовно летальними в залежності від ступеня модифікації фермента та його ролі в репродуктивному циклі вірусу. Генетичні взаємодії між вірусами можуть носити кооперативний і інтерферуючий характер.
Кооперативні взаємодії. Рекомбінації та перерозподіл генів між фрагментованими геномами призводить до перерозподілу генетичного матеріалу в популяціях. Вони відмічені по всіх групах ДНК-вмісних вірусів, у всіх РНК-вмісних вірусів з сегментованим геномом та у деяких РНК-вмісних вірусів з несегментованим геномом (поліовірус, вірус ящуру). Рівень рекомбінації дволанцюгових ДНК-вмісних вірусів є пропорційним розміру геномів. У РНК-вмісних вірусів при копіюванні “ + ” ланцюга в “ - ” ланцюг полімераза може “перескочити” з одного ланцюга на інший, створюючи гібрідну матрицю РНК Подібний механізм обумовлює появу генетичної мінливості у ВІЛ. Геном ВІЛ утворено двома ланцюгами “+” РНК, при транскрипції ДНК з РНК зворотна транскриптаза може “перескакувати” з одного ланцюга на інший. Якщо обидва ланцюга ідентичні, то подібне явище не призводить до наслідків, але при наявності двох вірусів-мутантів можлива поява рекомбінантів з іншими геномами. Високий рівень перерозподілу фрагментованими геномами спостерігається як у одно-, так и у дволанцюгових РНК-вірусів. Обмін фрагментами геномів у штамів вірусів грипу обумовлює появу нових типів поверхневих гемаглютининів і нейромінідаз у вірусів грипу (антигенний шифт). Функціональна взаємодія двох дефектних вірусів, за умов, коли кожен з них не може розмножуватись разом, забезпечує можливість їх сумісної реплікації і горизонтальної передачі. Слідуючу форму генетичних взаємодій складають фенотипичне змішування та фенотипічне маскування (псевдотипування). Фенотипичне змішування спостерігається при одночасовому зараженні клітини схожими за типом вірусами. В цьому випадку утворюються віріони з гібридними капсидами, які кодуються геномами двох вірусів (наприклад, поліо- та Коксакі-вірусів).
При фенотипичному маскуванні процес може развиватися і в зворотному напрямку при коінфікуванні вірусами ідентичного псевдотипу. Якщо віріони мають геном ІІ типа і заключені в капсид І типу, то дочірні популяції будуть включати капсид і геном ІІ типу, так як утворення всіх їх структурних компонентів кодує геном ІІ типу.
Интерференцією вірусів позначають стан нечутливості клітини, яка уже інфікована вірусом, до повторного зараження.
При гетерологічній інтерференції інфікування одним вірусом повністю блокує можливість реплікації іншого віруса в межах однієї клітини. Механізми гетерологічної інтерференції пов¢язані з блокуванням або руйнуванням специфічних клітинних рецепторів, або з придушенням трансляції будь-якої гетерологічної іРНК в інфікованій клітині.
При гомологічній інтерференції реплікація віруса з дефектним геномом можлива при сумісному зараженні з нормальним вірусом; в подібних взаємодіях останній визначається як вірус-помічник. Однак дефектний вірус може вмішуватися в його реплікативний цикл і утворювати дефектні інтерферуючі (ДІ) вірусні частки. ДІ-частки мають лише частину геному повного віруса, і не дивлячись на те, що у ДІ-вірусів експресуються деякі гени - їх основною властивостю є здатність до інтерференції з гомологічним вірусом. Циркулювання ДІ-часток і коінфекція з “нормальним” вірусом викликає явище в¢ялотекучої, тривалої формии захворювання. Взаємодія між вірусом, первинно заразившим клітину, і вірусом, вторинно прониклим до неї може реалізуватися двома шляхами: 1. Інфікована клітина може бути нечутливою до повторного зараження (первинне зараження може індукувати синтез інтерферонів,які інгибують реплікацію другого вірусу; у зараженій клітині може блокуватися синтез білків, які служать рецепторами для інших вірусів; можливе блокування трансляції іРНК другого віруса, обумовлене змінами 5¢-кінців іРНК; при репродукуванні первинного вірусу в зараженій клітині можуть формуватися дефекти регуляції експресії клітинних генів, які є необхідними для ранніх етапів реплікації другого віруса. 2. Репродукція віруса може зробити резистентну клітину чуттєвою до повторного зараження вірусом, що обумовлене здатністю першого віруса посилювати трансляцію іРНК другого віруса. Різноманітність вірусів багато в чому обумовлена змінами структури геномів; біологічну ефективність подібної мінливості визначають ряд факторів: зміни не повинні діяти на здатність вірусів репродукуватися в клітинах; вірус-мутант повинен мати перевагу перед “нормальним” вірусом. Найбільш перспективні мутації, що відводять вірус від захисних систем організма-хозяїна.
1.7.6. Репродукція вірусів
Загальні закономірності розмноження вірусів полягають в наступних положеннях: усі РНК-геномні віруси, окрім вірусу грипу і ретровірусів, розмножуються в цитоплазмі; усі віруси ДНК-вмісні, окрім вірусу віспи, розмножуються в ядрі; нуклеокапсидні вірусні білки синтезуються на вільних (не пов'язаних з клітинною мембраною) полірибосомах, а вільні білки - на мембранах шорсткої ендоплазматичної мережі; білки деяких вірусів піддаються каталітичному процесингу і глікозуванню; суперкапсидні білки глікозуються в процесі свого транспорту на зовнішню поверхню клітинної мембрани.
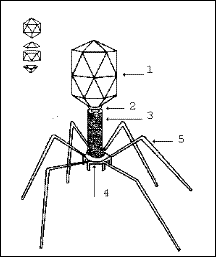
Рис. 17. Схема будови вірусу грипу
Схематично можна виділити два типи взаємодії вірусу і чутливої до нього клітини: продуктивний і інтеграційний. При продуктивному типі взаємодії вірусний геном функціонує в зараженій клітині більш-менш автономно, що проявляється в незалежній від клітинного геному репродукції генома вірусу. Цей тип взаємодії завершується утворенням нового покоління вірусних часток і загибеллю зараженої клітини. Таку реакцію на вірусну інфекцію називають літичною. У інших випадках клітина у якій розмножується вірус, може тривалий час зберігати свою життєздатність, а вірус при цьому видаляється з клітини шляхом брунькування.
При принципово іншому шляху взаємодії комплексу "вірус - клітина" відбувається об'єднання, інтеграція геномів цих двох організмів. Нова клітина з об'єднаним геномом може бути цілком життєздатною і дочірні клітини, що утворюються при діленні, також матимуть інтегроване клітинне і вірусне геноми. Одним з найважливіших проявів такої взаємодії вірусу і клітини є зміна деяких спадкових властивостей клітини. Якщо йдеться про взаємодію вірусу з бактеріаλльною клітиною, то такий окремий випадок носить назви лізогенія. Віруси, здатні викликати вірогенію (лізогенію), відносяться до групи помірних. Прикладом помірних вірусів є фаг лямбда. Між двома групами вірусів відсутня непереборна грань. У ряді випадків помірні віруси здатні до автономної репродукції в клітині або ж втрачають здатність до вірогенії в результаті мутації і набуває властивість типово вірулентних вірусів. У розмноженні вірусів виділяють ранні стадії, під час яких здійснюється підготовка вірусних структур до відтворення потомства, і пізні, включаючі синтез вірусспецифичних молекул, збірку віріонів і вихід вірусу з клітини. До ранніх стадій відносять адсорбцію вірусу на поверхні клітини, проникнення його в клітину і роздягання (депротеїнезацію) вірусу, що супроводжується відділенням захисних оболонок і деяких білків.
Адсорбція.
У основі адсорбції лежать два механізми: 1) неспецифічний, оборотний - віріони утримуються на поверхні клітини за допомогою електростатичних і Ван-дер-Вальсових сил; 2) специфічний, в основі якого лежить взаємодія специфічних рецепторів вірусу з відповідними рецепторами клітини. Клітинні рецептори різні для різних груп вірусів. Рецепторними функціями для ряду вірусів володіють глікопротеїди клітинних плазматичних мембран і суттєвим компонентом в них є вуглеводи. Рецептором для вірусів грипу є гли-копротеїд, що містить нейрамінову кислоту. Рецепторні функції мають також клітинні ліпіди. Клітина може бути заражена вірусом і у тому випадку, коли у неї відсутні специфічні вірусні рецептори (шляхом обробки клітини інфекційними вірусними нуклеїновими кислотами, наприклад, РНК вірусу поліомієліту; зараженні вірусами клітинних протопластів; використанні "гібридних" вірусних часток, що містять нуклеїнову кислоту одного вірусу, а білкову оболонку (тобто рецептори) іншого.
Рис. 18. Адсорбція бактеріофага Т4 на клітинній стінці кишкової палочки
На ефективність адсорбції вірусу на клітинній мембрані впливає концентрація вірусів, температура і стан клітини, наявність в середовищі електролітів (катіонів) і вільних амінокислот - кофакторів адсорбції, тканинна специфічність (рис. 18. 19).
Рис. 19. Адсорбція вірусів поліоми на мембрані чутливої клітини. Електронна мікроскопія. Комп'ютерна реконструкція.
Зазвичай тваринна клітина містить близько 500000 рецепторів і на клітині можуть сорбувати безліч віріонів. Проте, як правило, клітина стійка до повторного зараження вірусом того ж типу. Проникнення вірусу в клітину відбувається услід за адсорбцією і включає "роздягання", тобто депротеїнезацію віріонів і змінУ структури їх нуклеопротеїду. Проникнення віріона в клітину здійснюється у відповідність з тим же механізмом, що і проникнення в клітину позаклітинних поживних речовин, деяких гормонів, чинників зростання та ін., тобто шляхом рецепторного ендоцитоза. Механізм цього явища полягає в тому, що після прикріплення вірусів до клітинних рецепторів утворюються ендоцитарні вакуолі, які, зливаючись з іншими внутрішньоклітинними вакуолями, формують рецептосому - велику вакуоль з коротким періодом життя, пов'язану, що містить, з рецепторами вірусну частку.
Рис. 20. Адсорбція віріонів вірусу вісповакцини на поверхні культури клітин нирки зеленої мавпи (х24000).
У рецептосомі відбувається взаємодія поверхневих білків віріонів з ліпідами стінки вакуолі, що призводить до злиття ліпопротеїнової оболонки (пеплоса) оболонкових вірусів з плазмолемою і виходу внутрішнього компонента вірусів в цитоплазму. Цей механізм проникнення в клітину є універсальним як для багатьох оболонкових, так і для без оболонкових вірусів, що належать до різних таксономічних груп.
А Б Рис. 21. Послідовні етапи проникнення вірусу в клітини тварин і його "роздягання". А. Проникнення при частковому злитті оболонки вірусу і клітинної мембрани; Б. Проникнення вірусу з формуванням ендоцитозної вакуолі.
Взаємодія вірусної і клітинної мембран відбувається в більшості випадків при низьких (5,0 - 5,5) значеннях рН, що необхідно для придбання білками злиття необхідної конформації. Проникнення вірусів може здійснюватися через злиття мембран вірусу і клітини. Злиття обумовлене наявністю відповідних вірусних глікопротеїнів, внаслідок чого внутрішні структури вірусів опиняються в цитоплазмі заражених клітин, а вірусні оболонки - на поверхні клітин. Інший шлях проникнення реалізується у разі бактерій і вірусів бактерій - бактеріофагів. Цей шлях - проникнення в клітину тільки нуклеїнової кислоти вистачає добре вивчений на моделях Т-парних (Т-2, Т-4 і так далі) фагів кишкової палички. Фагова ДНК потрапляє в клітину в результаті скорочення білкового чохла відростка вірусу (рис. 17), чому передує адсорбція вірусу на клітинній стінці бактерії активація вірус-кодованого ферменту - лізоциму і часткове руйнування ним полісахаридного комплексу (рис. 22а, 22б).
1 2 3 4 Рис. 22а. Послідовні етапи інфікування бактеріальної клітини бактеріофагом групи Т-фагів кишкової палички 1. Адсорбція вірусу на клітинній поверхні; 2. Часткове руйнування полісахаридного комплексу клітинної стінки лізоцимом фага; 3. Введення білкової "голки" фага через мембрану клітини; 4. Інфікування клітини фагової ДНК
Роздягання. Для придбання інфекційної активності вірусна частка повинна лишитися ліпопротеїдної оболонки і частково - білків капсиду. Роздягання вірусу - багатоетапний процес, причому у деяких вірусів останні етапи роздягання проходять в ядрі зараженої клітини.
Рис. 22б. Проникнення ДНК фага Т2 в бактерію кишкової палички після адсорбції вірусу.
Пізні стадії. Інформація, записана у вірусному геномі, реалізується шляхом перемикання клітини на синтез вірусспецифічних молекул, що здійснюється двома способами: 1) утворенням коротких інформаційних РНК для індивідуальних білків (віруси герпесу, віспи, ВІЛ та ін.); 2) використанням вірусом власних геномів як інформаційних РНК з подальшою трансляцією гігантських поліпептидів - попередників функціонально активних білків віруси поліомієлиту, гепатиту та ін.). Перший спосіб характерний для вірусів ДНК-вмісних, а також для РНК- вмісних вірусів з негативним геномом і вірусів з фрагментованим геномом, у яких кожен фрагмент є окремим геном. Другий спосіб - для РНК- вмісних вірусів з позитивним геномом. У першому випадку інфекційний цикл в зараженій клітині починається з утворення іРНК, а в другому випадку - з синтезу білку. Поліпептид, що утворюється, - посередник розрізає клітинними і вірус-специфічними протеазами на функціонально активні. Слідує проте, що нарізування одного-двух білків-попередників або видалення невеликої ділянки поліпептиду відзначається при репродукції практично усіх вірусів і є необхідним для придбання ними інфекційної активності (цей процес називається протеолітичною активацією). У оболонкових РНК- вмісних вірусів нарізаються зазвичай поверхневі білки – глікопротеїди, що беруть участь в проникненні в клітину, тому протеолітична активація пов'язана із здатністю вірусу проникати в клітину. Важливою особливістю розмноження вірусів є те, що компоненти вірусу синтезуються в клітині окремо і лише потім з'єднуються в зрілу вірусну частку (диз’юктивний спосіб розмноження). Синтез вірусних нуклеїнових кислот і вірусних білків може відбуватися не одночасно і в різних частинах клітини. Передусім, після інфікування клітини вірусом настає різке пригнічення син-тезу клітинних нуклеїнових кислот і білку. Подальші етапи пов'язані з процесами транскрипції і трансляції вірусної нуклеїнової кислоти.
Транскріпція. Експресія вірусних геномів регулюється на рівні транскрипції, що призводить до утворення інформаційних РНК. Транскрипція вирусспецифічної інформації відбувається як за допомогою клітинних (більшість вірусів ДНК- вмісних), так і за допомогою вірусних ферментів-транскриптаз (віруси віспи, гепатиту В, грипу). Транскрипція РНК- вмісних вірусів здійснюється за участю транскриптаз, що кодуються вірусним геномом, які можуть бути як структурними, так і неструктурними білками. Наступна особливість полягає в тому, що синтетичний апарат еукаріотичних клітин не пристосований для трансляції поліцистронних іРНК. Віруси обходять цю перешкоду або шляхом синтезу іРНК, що включає декілька генів і що кодує великий "поліпротеїн", який потім розрізає на індивідуальні білки, або шляхом розподілу генетичної інформації по фрагментах генома, кожен з яких є окремим геном і транскрибується в окрему іРНК. У ДНК- вмісних вірусів іРНК синтезується в певній послідовності: спочатку транскрибуються надранні гени, потім ранні (ферменти, регулювальники - неструктурні білки), проміжні, і в останню чергу - пізні. Ферменти реплікації кодуються ранніми генами, структурні білки - пізніми. У паповавірусів ранні і пізні гени транскрибуються в різних напрямах на різних нитках кільцевої дволанцюжкової ДНК. Багато генів під час транскрипції перекривають один одного. Деякі ділянки ДНК прочитуються в різних рамках прочитування, внаслідок чого можуть утворитися два різні білкові продукти. Ці особливості транскрипції сприяють значній економії генетичного матеріалу і збільшенню місткості генетичної інформації вірусу. Для багатьох ДНК вірусів характерний синтез білков-трансактиваторів, що включають транскрипцію і стимулюючих експресію генів. Характерною особливістю вірусних трансактиваторів є відсутність специфічності: вони здатні активувати як експресію генів інших вірусів, так і генів клітин еукаріот. Одним із способів регуляції транскрипції є фосфорилювання і дефосфори-лювання білків, що впливає на зв'язування білків з нуклеїновою кислотою. Механізм реплікації забезпечує відтворення дочірніх вірусних геномів,які є точною копією материнських. Механізм реплікації у різних вірусів неоднаковий. У вірусів ДНК- вмісних в реплікації бере участь клітинна ДНК-полімераза. Реплікація кільцевої двохнитчастих ДНК починається з однієї точки і йде одночасно в обох напрямах з однаковою швидкістю. Як і у разі реплікації клітинної ДНК відбувається переривчастий синтез ініціаторів (приманок) - коротких ділянок РНК (РНК-праймерів) і коротких фрагментів ДНК (фрагменти Окадзаки), які потім ковалентно зв'язується в зростаючий ланцюжок. При утворенні лінійної дволанцюжкової ДНК (аденовіруси) вони синтезуються з обох кінців в напрямі від 51 - до 31-кінця, використовуючи вірусспецифічну ДНК-полімеразу. Реплікація вірусних РНК-геномів відбувається тільки за участю вірусспецифічних РНК-полімераз (тобто ферментів, синтез яких кодується вірусним геномом) і має потребу в утворенні антигенома - повнорозмірній дзеркальній копії комплементу вірусного генома:
РНК вірусна Þ РНК копія (антигеном) Þ РНК вірусна (дочірня) ¯ ¯ поліпротеїд РНК-поліимераза ¯ ¯¯¯¯¯(процесинг) ® ¯ Самозбмрання структурні віруса білки ®........... ®............ ®............
Одночасно на одній РНК матриці функціонує декілька репликаз і утворена структура називається реплікативним попередником. Приманкою для реплікації у пікорна- і калицивиірусов являється білок генома VРg, ковалентно пов'язаний з РНК(-) і РНК(+). Незначна маса вірусної нуклеїнової кислоти (106-107D) призводить до того, що місткості вірусної нуклеїнової кислоти недостатньо для запису інформації про синтез усіх необхідних при циклі вірусної реплікації білків. З цією метою віруси використовують ферментний апарат клітини-хазяїна, а у ряді випадків удаються до допомоги і інших вірусів. Наприклад, РНК вірусу-сателіта некрозу тютюну складається тільки з 1200 нуклеотидів, а кодована цією РНК білкова субодиниця включає 400 амінокислотних залишків. Очевидно, що ні для якої іншої інформації в геномі цього вірусу не вистачає місця, Тому вірус-сателліт некрозу тютюну здатний розмножуватися тільки в тих клітинах які одночасно заражені вірусом некрозу тютюну. Цікавим прикладом вірусів сателітів людини служить вірус гепатиту D, який репликується в гепатоцитах тільки у присутності вірусу сироваткового гепатиту В і використовує як капсид зовнішню білкову оболонку вірусу гепатиту В. Спільне розмноження вірусів гепатиту двох типів обумовлює особливо важка вдача інфекції з високою вірогідністю переходу в цироз печінки і в гепатокарциному.
Трансляція. Термін трансляція означає переведення генетичної інформації з матричної (інформаційної) РНК в послідовність амінокислотних залишків в білковому поліпептидному ланцюгу. Молекула іРНК через “шапочку" (кеп), розташовану на 5¢-кінці іРНК, зв'язується з малою рибосомальною субодиницею, яка рухається вздовж молекули РНК. По досягненні першого ініціюючого кодону АУГ з нею зв'язується велика рибосомальна субодиниця з транспортної РНК, і потім починається синтез білків. Трансляція триває доти, поки на рибосомі не з'явиться термінуючий кодон. Вірусні білки синтезуються на рибосомах, пов'язаних з клітинними мембранами; глікозування білків здійснюється клітинними ферментами і відбувається одночасно з синтезом поліпептиду. Синтезовані вірусні білки мігрують в різні ділянки клітини, наприклад в ядро, у випадку інфекції, обумовленої ядерними вірусами. Багато які вірусні білки модифікуються (глікозуються, ацетилюються, фосфорилюються, протеолітично нарізуються і т.д.) як в процесі трансляції, так і після неї.
Формування зрілих вірусних часток і вихід їх з клітини. Внаслідок синтезу вірусних білків і нуклеїнових кислот в клітині накопичується достатня їх кількість і створюються умови для формування зрілих вірусних часток. Найбільш простий механізм дозрівання віріонів у вірусів із спіральною симетрією, і вони утримують тільки один тип білків і нуклеїнову кислоту. Класичним представником таких вірусів вважається вірус тютюнової мозаїки (ВТМ). У ВТМ формування зрілої вірусної частки здійснюється шляхом самозбирання, тобто спонтанного об'єднання молекул в більш великі агрегати, в основі якого лежить специфічна взаємодія молекул білків і нуклеїнових кислот. У зв'язку з різноманіттям структури вірусів різноманітними є і способи формування віріонів. У складних вірусів спочатку формуються нуклеокапсид і серцевина, які потім взаємодіють з білками вірусних оболонок. Більшість оболонкових вірусів здобуває свої оболонки в ході відбруньковування через плазмолему або мембрану цитоплазматичної вакуолі. Таким чином, поліпротеїдна оболонка більшості вірусів є фрагментом плазмолеми клітини-господаря і за складом ліпідів дуже близька до неї. Виключенням є невідбруньковуючі віруси віспи і гепатиту В, які здобувають свої зовнішні поліпротеідні оболонки іншим способом.
Процес формування зрілих вірусних часток у складно влаштованих вірусів можна розглянути на прикладі Т-парних фагів кишкової палички. Морфогенез Т-парних фагів здійснюється через ряд послідовних стадій. При цьому три основних компоненти фагу - головка, відросток і фібрили (рис. 22) утворюються незалежно один від одного. У складних вірусів кожен подальший етап збирання віріону ініціюється завершенням попереднього етапу.
Рис. 23. Схематичне зображення віріону фага Т4 1 - головка з ДНК-геномом; 2 - комірець; 3 - нескорочений чохол відростка, що складається з 24 витків довгої білкової молекули; 4 - базальна пластинка зі шпичками, взаємодіючими з оболонкою бактерійної клітки; 5 - фібрили (нитки відростка фагу).
1. Утворюється головка фагу, що складається з більш ніж 10 типів білків і заповнюється довгою ниткою ДНК. Можливо, в цьому процесі беруть участь поліаміни (путресцин і спермідин), що входять до складу головки зрілого вірусу. 2. Затравкою для формування фагового відростка слугує базальна пластинка, побудована з 15 типів білків. 3. Після завершення збирання головки і відростка вони об'єднуються. 4. Нитки відростка, що складаються з продуктів 4 генів, збираються незалежно від інших вірусних структур, але прикріплюються вони до базальної пластинки тільки після прикріплення головки і відростка.
Лімітуючим фактором в збиранні вірусної частки є матриксний білок - гідрофобний білок, здатний до взаємодії як з білками вірусних оболонок, так і з ліпідами клітинної мембрани. Відразу ж після свого синтезу матриксний білок вбудовується в плазмолему з внутрішнього, цитоплазматичного боку ліпідного шару. Включення матриксного білка до плазмолеми є сигналом до збирання вірусної частки. Синтез матриксного білка має значні відмінності в клітинах різного походження і завдяки його лімітуючій ролі в збиранні вірусної частки, він є критичним фактором, що визначає тип інфекції. Незначна продукція матриксного білка призводить до формування абортивних інфекцій, що характеризуються відсутністю або незначною кількістю інфекційного потомства, і персистуючих інфекцій, при яких вірусспецифічні компоненти накопичуються в клітині і переходять до дочірніх клітин. Матриксний білок виявлено у вірусів грипу, сказу, ретровірусів.
Вихід вірусів з клітини здійснюється шляхом від брунькування, крізь пори клітинній мембрані (у простих вірусів) і шляхом “вибуху", при якому прості і безоболонкові віруси тварин і бактеріофаги виходять у позаклітинний простір після руйнування клітин. У руйнуванні бактерій під впливом фагів (рис. 23) бере участь фермент лізоцим, що розщеплює бактеріальні пептидоглікани. Віруси рослин і деякі віруси людини і тварин (віруси герпесу, респіраторно-синцитіальні віруси) можуть проникати в незаражену клітину з інфікованої через плазмодесми. Починаючи від моменту адсорбції на клітинній мембрані цикл розмноження вірусів займає від 20 - 40 хвилин (віруси бактерій) до 40-60 годин (віруси людини і тварин). Така велика різниця у часі, необхідному для вірусної репродукції, можливо пов'язана з відмінністю мікроорганізмів і клітин еукаріот і з різницею у відстані, яку повинні долати до моменту самозбирання складові елементи вірусів.
Якщо вірус передається від одного покоління клітин до іншого в ході клітинного поділу, латентний вірусний геном називається провірусом або, у разі вірусів бактерій, профагом, а клітини, які несуть такий профаг - лізогенними клітинами. Присутність геному вірусу в лізогенній культурі можна виявити при спонтанному вивільненні вірусу з клітинної популяції. У лізогенних клітинах не транскрибується жоден з вірусних генів, необхідних для вірусної інфекції. У експериментальних лізогенних культурах виявляється дуже невелика кількість вірусної іРНК, більша частина котрої синтезується на опероні, до складу якого входить ген, кодуючий синтез білків, що репресують транскрипцію решти вірусного геному. Інактивація репресора за допомогою опромінення ультрафіолетовими променями або обробки різними хімічними речовинами індукує розмноження вірусу.
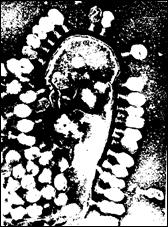
Рис 24. Частково лізована клітина Escherichia coli В, до поверхні якої приєднані віріони фагу Т4.
Явище лізогенії є одним з механізмів формування латентних вірусних інфекцій і лежить в основі перетворення нормальної клітки в пухлинну. Інтеграція вірусного геному в геном клітини і його подальше виключення зі складу клітинного геному створює умови для захоплення і перенесення (явище трансдукції) в нову клітину частини генетичного апарату клітину-господаря з інформацією, що не має безпосереднього відношення до життєвого циклу вірусу (явище конверсії). У ході конверсії, наприклад, бактерії роду Salmonella здобувають нові полісахаридні антигени в складі клітинної оболонки, а здатність збудників дифтерії (бактерій Corinebacterium diphtheriae) продукувати токсин залежить від бактеріофагу, що знаходиться в бактеріальній клітині в стані профагу.
|
|||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 164; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.148.124 (0.069 с.) |



 Рецептори служать не лише для прикріплення вірусів до клітинної поверхні але і для їх подальшого транспортування усередині клітини. Вірусні рецептори можуть бути унікальними органелами, видимими в електронному мікроскопі (хвостові структури у Т-фагів (мал. 11), грибоподібні вирости оболонки вірусу імунодефіциту людини) або морфологічно менш виражені структури, що складаються з глікопротеїдів на поверхні вірусних оболонок (гемаглютинін у ортоміксовірусів). Рецепторні ділянки зазвичай розташовуються на дні поглиблень і щілин на поверхні віріона, що таким чином захищає їх від блокуючої дії специфічних противірусних антитіл, діаметр яких більше діаметру щілини.
Рецептори служать не лише для прикріплення вірусів до клітинної поверхні але і для їх подальшого транспортування усередині клітини. Вірусні рецептори можуть бути унікальними органелами, видимими в електронному мікроскопі (хвостові структури у Т-фагів (мал. 11), грибоподібні вирости оболонки вірусу імунодефіциту людини) або морфологічно менш виражені структури, що складаються з глікопротеїдів на поверхні вірусних оболонок (гемаглютинін у ортоміксовірусів). Рецепторні ділянки зазвичай розташовуються на дні поглиблень і щілин на поверхні віріона, що таким чином захищає їх від блокуючої дії специфічних противірусних антитіл, діаметр яких більше діаметру щілини.