Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Мембрано-опосредованное вз-действие гормоновСодержание книги
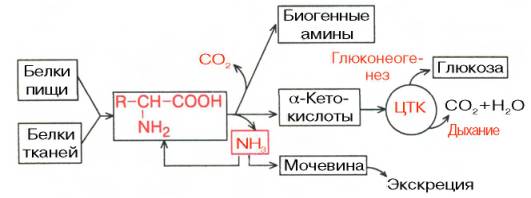
Поиск на нашем сайте Катаболизм АМК Промежуточный метаболизм АМК белковых мол-л, как и других питательных в-в в живых орг-мах, включает катаболические (распад до конечных продуктов обмена), анаболические (биосинтез АМК) процессы, а также ряд других специфических превращений, сопровождающихся обр-нием биологически активных соединений. Условно промежуточный метаболизм АМК можно разделить на общие пути обмена и индивидуальные превращения отдельных АМК.
Рис. 12.2. Катаболизм АМК.
Билет 27 Стеролы и стероиды СТЕРИНЫ (стеролы), алициклич. прир. спирты, относящиеся к стероидам; составная часть неомыляемой фракции животных и растит. липидов. Стерины присутствуют практически во всех тканях животных и растений и явл наиб. распространенными представителями стероидов в природе. В зависимости от источника подразделяются на животные (зоостерины), растительные (фитостерины), стерины грибов (микостерины) и микроорг-мов. Такая классификация, однако, имеет ограничения для морских орг-мов, отличающихся исключительным разнообразием стр-р и состава стеринов. Основной стерин высших животных - холестерин широко распростр и у др. орг-мов. Хим. св-ва стеринов мало отличаются от св-в алициклич. спиртов и алкенов. Особенность стероидной стр-ры стеринов проявл, напр., в их способности образовывать прочные труднорастворимые комплексы с дигитонином и нек-рыми др. стероидными сапонинами, что широко использовалось для выделения и очистки холестерина в ранний период исследований. Стр-ра и св-ва соед., содержащих двойные связи у атомов С-5 и С-22 ( 5,22-С), благоприятны для использования их в качестве исходных соед. в хим. синтезе ряда прир. стероидов и их аналогов, в т.ч. гормонов млекопитающих, насекомых и растений. Биосинтез стеринов, как и др. стероидов, осуществляется из уксусной и мевалоновой к-т в результате многостадийного ф-тативного процесса через непосредственный предшественник - сквален, циклизация к-рого в орг-мах животных и растений протекает по-разному. Превращение этих соед. в другие стерины-сложный и не до конца изученный процесс, протекающий в разл. биол. объектах неодинаково. Одно из отличий связано с присутствием у растений биосинтетич. механизмов алкилирования, к-рых нет у животных, в результате чего у растений преобладают стерины С28-С29, у животных-С27. Сами стерины, в свою очередь,-предшественники др. стероидов в живых орг-мах: у животных-желчных к-т, стероидных гормонов, вит-а D, у растений - агликонных частей стероидных гликозидов, нек-рых регуляторов роста растений и др. Большинство биосинтетич. превращений стеринов имеет окислит. хар-р и протекает под-действием оксидоредуктаз, среди к-рых важная роль принадлежит цитохромам Р-450. В природе стерины- биосинтетич. предшественники мн. стероидных биорегуляторов, осн. стр-рные компоненты (наряду с белками и фосфолипидами) клеточных мембран. Предполагают, что они выполняют при этом не только пассивную (стр-рную) ф-цию, но и влияют на клеточный метаболизм. Свои ф-ции в орг-ме млекопитающих стерины реализуют в виде комплексов с белками (липопротеидов) и сложных эфиров высших жирных к-т, являясь их переносчиками во все органы и ткани через систему кровотока. Фитостерины, напр. -ситостерин, в отличие от холестерина не усваиваются орг-мом человека. Большое разнообразие стеринов у растений, дрожжей и беспозвоночных, резко отличающееся от стеринов животных и человека, не имеет объяснения с функцион. точки зрения. Анализ стеринов основан на сочетании хроматографич. и спектральных методов; весьма эффективна хромато-масс-спект-рометрия, позволяющая в процессе анализа фракции идентифицировать ее по типу фрагментации. Известные цветные р-ции нек-рых типов стеринов представляют в настоящее время лишь теоретич. интерес. В пром-сти стерины выделяют из животного или растит. стеринсодержащего сырья: холестерин-из продуктов мясопереработки (мозга крупного рогатого скота), стиг-мастерин-из масла соевых бобов, -ситостерин-из отходов целлюлозно-бум. производства, эргостерин-из дрожжей. Применяют стерины гл. обр. для получения стероидных гормо-нов и производных на их основе, а также вит-а D и др. фармацевтич. препаратов. СТЕРОИДЫ, группа биологически важных прир. соед., в основе стр-ры к-рых лежит скелет циклопентанопер-гидрофенантрена (гонана, стерана; ф-ла I). Входят в состав всех растит. и животных орг-мов.
По номенклатуре ИЮПАК полная нумерация в стероидах производится так, как показано в ф-ле П. Если один или более атомов С отсутствуют, нумерация оставшейся части сохраняется. В формулах стероидов связи атомов или групп, располагающихся за плоскостью кольцевой системы (-конфигурация), изображают пунктирной линией, атомы и группы, располагающиеся перед плоскостью (-конфигурация), -жирной линией; связи с неопределенной конфигурацией обозначают волнистой линией. Назв. частично ненасыщенных стероидов производят от назв. насыщенных посредством окончания "ен". В названиях стероидов, содержащих внутри стр-ры трехчленное кольцо, вводится префикс "цикло" с цифровым указанием положения и стереохимией обозначения; общее назв. таких стероидов-циклостероиды. Природные стероиды имеют транс- или цис-сочленение колец А и В, транс-сочленение колец В и С и обычно транс-сочленекие С и D. Циклогексановые кольца, как правило, находятся в энергетически выгодной конформации кресла. Большинство стероидов содержат гл. обр. группы СН3 у атомов С-10 и С-13, группу ОН (или кетогруппу) у атома С-3 и боковую цепь у атома С-17, содержащую до 8-10 атомов углерода. Стероиды включают в себя стерты (стеролы), сапонины, стероидные алкалоиды, стероидные гормоны, желчные к-ты, желчные спирты, гликозиды сердечные, вит- D. Все стероиды (подобно др. изопреноидам) объединяются одной схемой биогенеза: в живой клетке из уксусной k-ты через ацетилкоф-т А и мевалоновую к-ту строятся изопрено-вые фрагменты -изопентенилпирофосфат и изомерный ему диметилаллилпирофосфат, к-рые путем конденсации обр-ют общий биогенетич. предшественник -сквален; эпоксид последнего при циклизации с послед. окислит. отщеплением неск. углеродных атомов превращается в стероид. Стероиды выделяют из спинного мозга и желчи рогатого скота, из щелочного гидролизата дрожжей, растит. масел и животных жиров, отходов целлюлозно-бумажной пром-сти, синтезируют из неприродного сырья. Полный синтез осуществлен для мн. природных стероидов-холестерина, андрогенов, гестагенов, кортикостероидов, эстрогенов и др. Стероиды - биол. регуляторы. Способность к биосинтезу стероидов наиб. выражена у высших позвоночных. Насекомые стероиды не вырабатывают, а получают с пищей, однако ф-ция их линьки контролируется стероидами. Применяют стероиды гл. обр. в медицине (лек. ср-ва, гормоны и др.). Лактоза и ее св-ва Лактоза, или млочный сахар (бэта-D-галактопиранозил-(1-->4) – альфа-D-глюкопираноза), - основной дисахарид молока, сожержит свободный полуацетальный гидроксид при С-1 и обладает восстанавливающими св-вами. Олигосахариды группы лактозы представляют собой продукты гликозилирования мол-лы лактозы остатками моносахаридов (L-фукозы, N-ацетил-D-глюкозамина, D-галактозы, N-ацетилнейраминовой к-ты), т.е. трисахариды. Одна из ф-ций олигосахаридов группы лактозы - формирование бактериальной флоры в кишечнике новорожденных, необходимой для нормального пищеварения. Лигазы ТРНК
КоА Ацетил КоА В кишечной стенки всосавшиеся ацилгицерины распадаются под действием тканевых липаз с образованием свободных жирных кислот и глицерола Часть моноацилглицеринов может превращаться в триащгаглицерины без предварительного расшепления йо так называемый моноацклглицериновый путь ресинтеза Все высшие жирные кислоты всосавшиеся к, кишечника используются в энтероцитах для ресинтеза различных лилидов Но перед тем как: вклю иться в различные липиды высшие жирные кислоты должны быть активированы Процесс активации высших жирных кислот состоит из 2 этапов 1 этап. За счет взаимодействия жирных кислот и использования специального фермента образуется ацшюденилаты (так называемый термодинамический контроль направления процесса) ЖК + АТФ -> R-С-АМФ + пирофосфат расщ. до ФК.(термодинамический контроль) 2 этап Происходит образование активной жирной кислоты соединенной с КоА и высвобождение АМФ Образование ацилКоА катализируется специальным ферментом, причем он катализирует и первую и вторую реакцию ацилКо Чсиптетата (тиокиназа) R-С-АМФ + НSКоА -> R-С-SКоА + АМФ В ходе активации высшей жирной кислоты АТФ распадается до АМФ и 2 остатков фосфорной кислоты таким жирные кислоты участвуют в активированной форме. 6. Биосинтез пуриновых нуклеотидов
|
||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 330; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.11 (0.009 с.) |