
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Тема 3. 8. Механизмы, обеспечивающие разнообразие белков у эукариотов
У эукариотов набор и количество синтезирующихся белков может зависеть от доступности генов для транскрипции, перестройки генов, альтернативного сплайсинга и изменения стабильности мРНК. 1. В организме человека имеется более 200 различных типов клеток, существенно различающихся по структуре и функциям, хотя количество и структура ДНК в них практически одинаковы. Существование специализированных органов и тканей зависит от экспрессии генов, при которой в дифференцированных клетках разных тканей транскрибируются разные участки хроматина. Это достигается благодаря существованию механизма стойкой репрессии транскрипции части генов на протяжении всей жизни организма. В разных тканях стабильной репрессии подвергаются разные гены. Формирование зон стабильной репрессии происходит в процессе онтогенеза и дифференцировки клеток. Транскрипционно неактивные или стабильно репрессированные участки хроматина получили название гетерохроматина. Стойкая репрессия генов в участках гетерохроматина обеспечивается: • высококонденсированным состоянием ДНК; • метилированием дезоксицитидина в CpG-последовательностях ДНК (эта модификация изменяет конформацию хроматина и препятствует транскрипции); • связыванием ДНК с гистонами и образованием нуклеосом. В хроматине клеток разных органов и тканей наряду с нетранскрибируемыми зонами ДНК имеются активные или потенциально активные участки - участки эухроматина. В областях эухроматина на ДНК расположены транскрибируемые гены. Эти области характеризуются: • наличием молекул гистонов, в которых метилированы или ацетилированы аминогруппы в радикалах Лиз и Арг, а остаток Сер фосфорилирован. Это снижает суммарный положительный заряд гистонов и ослабляет их связь с ДНК в нуклеосомах; • наличием участков, более чувствительных к действию ДНКаз. За исключением лимфоцитов каждая клетка организма содержит один и тот же набор генов. Однако «активность» одних и тех же генов в клетках тканей может существенно различаться. Например, в ретикулоцитах участок ДНК, содержащий ген β-глобина, находится в области «активного» хроматина, а в мышечных клетках локализован в районе «неактивного» хроматина. 2. Перестройка генов. Это явление наиболее отчетливо наблюдается при формировании разнообразия иммуноглобулинов (Ig). В зародышевых и всех соматических клетках нет полных генов, кодирующих L- и H-цепи Ig. Информация об отдельных участках цепи Ig представлена в виде соответствующих фрагментов молекулы ДНК- сегментов. В молекулах ДНК сегменты, кодирующие вариабельные (V) и константные (C) домены L- и Н-цепей (легких и тяжелых цепей соответственно), разделены протяженными нуклеотидными последовательностями. Обнаружено три разных семейства сегментов. Это два семейства, ответственные за синтез легких цепей, - λ (локализованное в 22-й хромосоме) и κ (2-я хромосома) - и одно семейство, содержащее информацию о всем разнообразии Н-цепей (14-я хромосома). Каждая легкая цепь кодируется тремя отдельными сегментами: V (вариабельным), J (соединяющим) и С (константным). Для κ-цепей существует по 250 V-сегментов, 5 J-сегментов и 1 С-сегмент (рис. 3.24). Тяжелые цепи кодируются 4 сегментами ДНК: VH, D (сегмент разнообразия), JH и CH. У человека около 1000 Vjj-сегментов, более 12 D-сегментов и 4 Jjj-сегмента.
В ходе дифференцировки клеток - предшественников В-лимфоцитов полные гены L-цепей собираются из трех сегментов в результате одной соматической рекомбинации: один из 250-ти VL сегментов соединяется с одним из 5 J-сегментов с образованием смешанного VJ-экзона,
Рис. 3.24. Синтез легкой κ-цепи иммуноглобулина Рекомбинация объединяет один V-сегмент вариабельного домена с одним из J-сегментов и приближает их к константному сегменту (C). После транскрипции образуется первичный транскрипт (пре-мРНК). Удаление лишних J-сегментов и интронов происходит в ходе посттранскрипционной модификации этой пре-мРНК, что приводит к появлению зрелой мРНК, после трансляции которой появляется белковый вариант легкой цепи Ig. Молекула ДНК показана в виде одной цепи а нуклеотидная последовательность ДНК между выбранными сегментами удаляется. С помощью такой перегруппировки VL- и JL-сегменты, выбранные из множества им подобных, транскрибируются с образованием единой молекулы мРНК предшественника. После созревания она превращается в мРНК, кодирующую одну из L-цепей Ig. Сходным образом образуются легкие λ-цепи.
При формировании полного гена Н-цепи первоначально собирается смешанный экзон из двух любых D- и J-сегментов в процессе первой рекомбинации. Затем в ходе второй рекомбинации к DJ-экзону присоединяется один из VH-сегментов и образуется полный ген Н-цепи IgM, поскольку к смешанному VHDHJH-экзону, кодирующему вариабельный домен Н-цепи, ближе всего находится СH-сегмент. Десять СН-сегментов константной области: Сμ Сδ, Су3, Су1, Са1, Су2, Су4, Се и др. - содержат информацию о строении доменов этой области и определяют классы и подклассы иммуноглобулинов IgM, IgG, IgA и т.д. Соединяя различные кодирующие V-, D-, J- и C-сегменты, организм может синтезировать миллионы разных молекул Ig. Иммуноглобулины (Ig) являются гликопротеинами, поэтому одной из посттрансляционных модификаций, которым подвергаются Н-цепи Ig, является синтез олигосахарида в С-области молекулы. Первыми в иммунном ответе появляются IgM, поскольку ген, кодирующий Сμсегмент Н-цепи, расположен на 5'-конце кодирующей области ДНК впереди всех остальных С-сегментов. Переключение классов Ig сопряжено с дополнительной специфической рекомбинацией, в процессе которой удаляются С-сегменты между полным геном вариабельной части и С-областью синтезируемого класса Ig. Таким образом, перестройка генетического материала в процессе формирования полных генов Ig проходит в несколько последовательных этапов, и каждый из них приурочен к строго определенной стадии дифференцировки В-лимфоцитов. Аналогичные процессы наблюдаются и в ходе дифференцировки Т-лимфоцитов. 3. Альтернативный сплайсинг мРНК. В ядре синтезируется значительно больший набор первичных транскриптов, чем количество зрелых мРНК в цитоплазме. Контрольные механизмы клетки определяют, какие из пре-мРНК подлежат процессингу, а какие будут расщепляться до нуклеотидов. Ранее уже говорилось о том, что созревание мРНК включает несколько этапов: -кэпирование 5'-конца; - присоединение поли-(А)-фрагмента к 3'-концу; - сплайсинг (удаление интронов). В некоторых случаях наблюдается альтернативный сплайсинг в сочетании с полиаденилированием, что приводит к образованию нескольких «зрелых» мРНК, кодирующих различающиеся между собой белки из одного и того же первичного транскрипта. Так, в парафолликулярных клетках щитовидной железы в ходе транскрипции гена кальцитонина образуется мРНК, которая содержит информацию о гормоне белковой природы, ответственном за регуляцию обмена ионов кальция (рис. 3.25). В мозге тот же первичный транскрипт подвергается другому варианту сплайсинга и полиаденилирования, и получается мРНК, кодирующая белок, ответственный за вкусовое восприятие.
Альтернативный сплайсинг описан для большого числа транскрибируемых генов. В частности, он показан для генов, кодирующих Ig. На ранних стадиях развития пре-В-лимфоциты продуцируют IgM, которые связаны с клеточной мембраной. Они синтезируются на мРНК, содержащей экзон, в котором имеется информация о гидрофобном участке С-области Ig. С помощью этого участка происходит «заякоревание» IgM в мембране (рис. 3.26).
Участок ДНК Сμсегмента (показана одна цепь ДНК) включает экзоны и содержит два стоп-кодона: один - в области интрона между двумя экзонами, второй - в составе второго экзона. В процессе транскрипции этого участка ДНК образуются два варианта первичного транскрипта как результат прочтения разных стоп-кодонов. После трансляции транскриптов образуются две разные формы IgM: «заякоренной» (1) в мембране и секреторной (2) В ходе дифференцировки образуются плазматические клетки, в которых за счет остановки транскрипции на стоп-кодоне 1, входящем в интрон, и альтернативного полиаденилирования исчезает экзон, кодирующий гидрофобный участок молекулы, и синтезируются укороченные молекулы антител, которые секретируются в кровь.
|
|||||||||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 1016; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.217.84.171 (0.011 с.) |
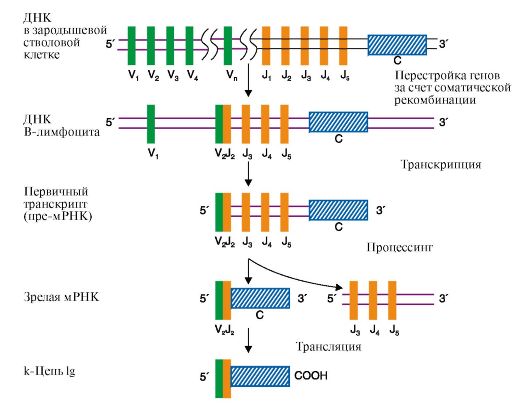
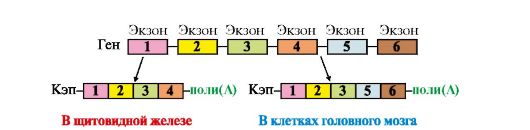 Рис. 3.25. Сплайсинг пре-мРНК гена кальцитонина в клетках щитовидной железы и мозга
Рис. 3.25. Сплайсинг пре-мРНК гена кальцитонина в клетках щитовидной железы и мозга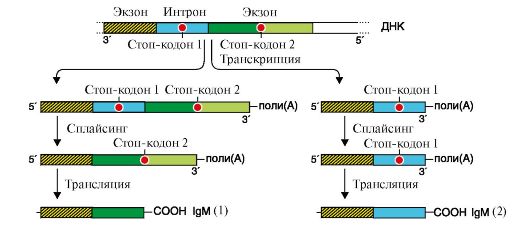 Рис. 3.26. Образование матриц для синтеза мембранно-связанных (1) и секреторных (2) форм Ig.
Рис. 3.26. Образование матриц для синтеза мембранно-связанных (1) и секреторных (2) форм Ig.


