Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Прямое окислительное дезаминирование.Стр 1 из 9Следующая ⇒
Прямое окислительное дезаминирование. Этот путь дезаминирования преобладает в мышцах при интенсивной работе, в результате которой накапливается молочная кислота. Выделяющийся аммиак предотвращает закисление среды в клетках, вызванное образованием лактата. 1). Анаэробное прямое окислительное дезаминирование существует только для глутаминовой кислоты, катализируется только глутаматдегидрогеназой, превращает глутамат в α-кетоглутарат. Этот тип дезаминирования теснейшим образом связан с трансаминированием аминокислот и формирует с ним процесс трансдезаминирования.
Реакция прямого окислительного дезаминирования глутаминовой кислоты. Окислительное дезаминирование глутамата – обратимая реакция и при повышении концентрации аммиака в клетке может протекать в боратном направлении, как восстановительное аминирование α-кетоглутарата. Глутаматдегидрогенназа очень активна в митохондриях клеток практически всех органов кроме мышц. Этот фермент – олигомер, состоящий из 6 субъединиц (молекулярная масса 312 кД). Глутаматдегидрогеназа играет важную роль, так как являктся регуляторным ферментом аминокислотного обмена. Аллостерические ингибиторы глутаматдегидрогеназы (АТФ, ГТФ, NADH) вызывают диссоциацию фермента и потерю глутаматдегидрогеназой активности. Высокие концентрации АДФ активируют фермент. Таким образом низкий энергитический уровень в клетках стимулирует разрушение аминокислот и образование α-кетоглутарата, поступающего в ЦТК как энергетический субстрат. Глутаматдегидрогеназа может индуцироваться стероидными гормонами (кортизолом). 2).Аэробное прямое окислительное дезаминирование катализируется оксидазами D– аминокислот (D–оксидазы) в качестве кофермента использующими ФАД, и оксидазами L-аминокислот (L-оксидазы) с коферментом ФМН. В организме человека эти ферменты присутствуют, но практически не активны.
Реакция, катализируемая оксидазами D- и L- аминокислот Оксидаза L-аминокислот В печени и почках обнаружен фермент оксидаза L-аминокислот, способный дезаминировать некоторые L-аминокислоты. (схема 2)
Коферментом в данной реакции выступает FMN. Однако вклад оксидазы L-аминокислот в дезаминирование, очевидно, незначителен, так как оптимум её действия лежит в щелочной среде (рН 10,0). В клетках, где рН среды близок к нейтральному, активность фермента очень низка.
Оксидаза D- аминокислот обнаружена в почках и печени. Это FAD- зависимый фермент. Оптимум рН оксидазы лежит в нейтральной среде, поэтому фермент более активен, чем оксидаза L-аминокислот. Роль оксидазы D- аминокислот способствует их превращению в соответствующие L- изомеры. (рисунок 2) 1. Непрямое дезаминирование (трансдезаминирование). Большинство аминокислот не способно дезаминироваться в одну стадию, подобно Глу. Аминогруппы таких аминокислот в результате трансаминирования переносятся на α-кетоглутарат с образованием глутаминовой кислоты, которая затем подвергается прямому окислительлному дезаминированию. Такой механизм дезаминирования аминокислот в 2 стадии получил название трансдезаминирования, или непрямого дезаминирования:
Первая стадия заключается в обратимом переносе NH2 – группы с аминокислот на кетокислоту с образованием новой кетокислоты – этот перенос называется трансаминирование и его механизм сложен. В качестве кетокислоты – акцептора («кетокислота 2») в организме обычно используется α-кетоглутаровая кислота, которая превращается в глутамат («аминокислота 2»). В результате трансаминирования свободные аминокислоты теряют α- NH2 – группы и превращаются в соответствующие кетокислоты. Далее их кетоскелет катаболизируется специфическими путями и вовлекается в цикл трикарбоновых кислот и тканевое дыхание, где сгорает до CO2 и H 2O. При необходимости (например, голодание) углеродный скелет глюкогенных аминокислот может использоваться для синтеза глюкозы в глюконеогенезе. Вторая стадия состоит в отщЕплении аминогруппы от аминокислоты 2 – дезаминирование. В организме человека дезаминирование подвергается только глутаминивая кислота. Вторая стадия осуществляется глутаматдегилрогеназой, которая имеется в митохондриях всех клеток, кроме мышечных. Учитывая тесную связь обоих этапов, непрямое окислительное дезаминирование называют трансдезаминированием.
ВИТАМИН А.
Витамин А (ретинол) - циклический, ненасыщенный, одноатомный спирт. Источники. Витамин А содержится только в животных продуктах: печени крупного рогатого скота и свиней, яичном желтке, молочных продуктах; особенно богат этим витамином рыбий жир. В растительных продуктах (морковь, томаты, перец, салат и др.) содержатся каротиноиды, являющиеся провитаминами А. В слизистой оболочке кишечника и клетках печени содержится специфический фермент каротиндиоксигеназа, превращающий каротиноиды в активную форму витамина А. Суточная потребность витамина А взрослого человека составляет от 1 до 2,5 мг витамина или от 2 до 5 мг р-каротинов. Обычно активность витамина А в пищевых продуктах выражается в международных единицах; одна международная единица (ME) витамина А эквивалентна 0,6 мкг β-каротина и 0,3 мкг витамина А. Биологические функции витамина А. В организме ретинол превращается в ретиналь и ретиноевую кислоту, участвующие в регуляции ряда функций (в росте и дифференцировке клеток); они также составляют фотохимическую основу акта зрения. Наиболее детально изучено участие витамина А в зрительном акте (рис. 3-3). Светочувствительный аппарат глаза - сетчатка. Падающий на сетчатку свет адсорбируется и трансформируется пигментами сетчатки в другую форму энергии. У человека сетчатка содержит 2 типа рецепторных клеток: палочки и колбочки. Первые реагируют на слабое (сумеречное) освещение, а колбочки - на хорошее освещение (дневное зрение). Палочки содержат зрительный пигмент родопсин, а колбочки - йодопсин. Оба пигмента - сложные белки, отличающиеся своей белковой частью. В качестве кофермента оба белка содержат 11-цисретиналь, альдегидное производное витамина А. Ретиноевая кислота, подобно стероидным гормонам, взаимодействует с рецепторами в ядре клеток-мишеней. Образовавшийся комплекс связывается с определёнными участками ДНК и стимулирует транскрипцию генов (см. раздел 4). Белки, образующиеся в результате стимуляции генов под влиянием ретиноевой кислоты, влияют на рост, дифференцировку, репродукцию и эмбриональное развитие (рис. 3-4). Основные клинические проявления гиповитаминоза А. Наиболее ранний и характерный признак недостаточности витамина А у людей и экспериментальных животных - нарушение сумеречного зрения (гемералопия, или "куриная" слепота). Специфично для авитаминоза А поражение глазного яблока - ксерофтальмия, т.е. развитие сухости роговой оболочки глаза как следствие закупорки слёзного канала в связи с ороговением эпителия. Это, в свою очередь, приводит к развитию конъюнктивита, отёку, изъязвлению и размягчению роговой оболочки, т.е. к кера-томаляции. Ксерофтальмия и кератомаляция при отсутствии соответствующего лечения могут привести к полной потере зрения. БЕЛКИ ПЛАЗМЫ КРОВИ. альбумин, α1-, α2-, β- и γ-глобулины. Разделение на альбумин и глобулин первоначально основывалось на различии в растворимости: альбумины растворимы в чистой воде, а глобулины — только в присутствии солей. Синтез белков плазмы крови осуществляется преимущественно в клетках печени и ретикулоэндотелиальной системы. Физиологическая роль белков плазмы крови многогранна. 1. Белки поддерживают коллоидно-осмотическое (онкотическое) давление и тем самым постоянный объем крови. Содержание белков в плазме значительно выше, чем в тканевой жидкости. Белки, являясь коллоидами, связываютводу и задерживают ее, не позволяя выходить из кровяного русла. Несмотря на то что онкотическое давлениесоставляет лишь небольшую часть (около 0,5%) от общего осмотического давления, именно оно обусловливает преобладание осмотического давления крови над осмотическим давлением тканевой жидкости. Известно, что в артериальной части капилляров в результате гидростатического давления безбелковая жидкость крови проникает в тканевое пространство. Это происходит до определенного момента – «поворотного», когда падающее гидростатическое давление становится равным коллоидно-осмотическому. После «поворотного» момента в венозной части капилляров происходит обратный ток жидкости из ткани, так как гидростатическое давление стало меньше, чем коллоидно-осмотическое. При иных условиях в результате гидростатического давления в кровеносной системе водапросачивалась бы в ткани, что вызвало бы отек различных органов и подкожной клетчатки.
2. Белки плазмы принимают активное участие в свертывании крови. Ряд белков, в том числе фибриноген, являются основными компонентами системы свертывания крови. 3. Белки плазмы в известной мере определяют вязкость крови, которая, как отмечалось, в 4–5 раз выше вязкостиводы и играет важную роль в поддержании гемодинамических отношений в кровеносной системе. 4. Белки плазмы принимают участие в поддержании постоянного рН крови, так как составляют одну из важнейшихбуферных систем крови. 5. Важна также транспортная функция белков плазмы крови: соединяясь с рядом веществ (холестерин, билирубин и др.), а также с лекарственными средствами (пенициллин, салицилаты и др.), они переносят их к тканям. 6. Белки плазмы играют важную роль в процессах иммунитета (особенно это касается иммуноглобулинов). 7. В результате образования с белками плазмы недиализируемых комплексов поддерживается уровень катионов вкрови. Например, 40–50% кальция сыворотки связано с белками, значительная часть железа, магния, меди и других элементов также связана с белками сыворотки. 8. Наконец, белки плазмы крови могут служить резервом аминокислот. Современные физико-химические методы позволили открыть и описать около 100 различных белковых компонентов плазмы крови. Особое значение приобрело электрофоретическое разделение белков плазмы (сыворотки) крови.
Пониженное количество белка (гипопротеинемия) возникает по причине: недостаточного поступления белка в организм - результат длительного голодания, безбелковой диеты, нарушения деятельности желудочно-кишечного тракта; повышенной потери белка - в результате острых и хронических кровотечениях, злокачественных новообразованиях; нарушения образования белка - как результат недостаточности функции печени (гепатит, цирроз, дистрофия печени). Повышенное количество белка (гиперпротеинемия) возникает по причине потери части внутрисосудистой жидкости - при перегревании организма, обширных ожогах, тяжелых травмах, холере, миеломной болезни Белковый состав плазмы крови очень разнообразен. Современная медицина идентифицировала более 100 различных белков плазмы. Наиболее простые белки - альбумины, глобулины и фибриноген находятся в плазме в больших количествах, остальные - в ничтожно малых. По форме и величине молекул белки крови разделяют на альбумины и глобулины. Наиболее распространенный белок плазмы крови — альбумин (более 50% всех белков, 40-50 г/л). Они выступают как транспортные белки для некоторых гормонов, свободных жирных кислот, билирубина, различных ионов и лекарственных препаратов, поддерживают постоянство коллоидно-осмотического постоянства крови, участвуют в ряде обменных процессов в организме. Синтез альбумина происходит в печени. 16. ЛИПОПРОТЕИНЫ В ТЕТРАДИ Д БЫТЬ ВИТ В1 Витамин В1 (тиамин; антиневритный), как отмечалось, был первым кристаллическим витамином, выделенным К. Функом в 1912 г. Позже был осуществлен его химический синтез. Наряду с аминогруппой витамин B1 содержит атомысеры, поэтому он был назван тиамином. В химической структуре его содержатся два кольца – пиримидиновое и тиазоловое, соединенных метиленовой связью. Обе кольцевые системы синтезируются отдельно в виде фосфорилированных форм, затем объединяются через четвертичный атом азота.
Тиамин хорошо растворим в воде. Водные растворы тиамина в кислой среде выдерживают нагревание до высокихтемператур без снижения биологической активности. В нейтральной и особенно в щелочной среде витамин B1, наоборот, быстро разрушается при нагревании. Этим объясняется частичное или даже полное разрушение тиаминапри кулинарной обработке пищи, например выпечке теста с добавлением гидрокарбоната натрия или карбоната аммония. При окислении тиамина образуется тио-хром, дающий синюю флюоресценцию при УФ-облучении. На этом свойстве тиамина основано его количественное определение. Витамин B1легко всасывается в кишечнике, но не накапливается в тканях и не обладает токсическими свойствами. Избыток пищевого тиамина быстро выводится с мочой. В превращении витамина B1в его активную форму – тиаминпирофосфат (ТПФ), называемый также тиамин-дифосфатом (ТДФ), участвует специфический АТФ-зависимыйфермент тиаминпирофосфокиназа, содержащаяся главным образом в печени и ткани мозга. Опытами с меченным32Р АТФ доказан перенос на тиамин целиком пирофосфатной группы в присутствии фермента. ТПФ имеет следующее строение:
Если витамин B1поступает с пищей в виде ТПФ, то пирофосфатная группа отщепляется от него под действием кишечных пирофосфатаз. При отсутствии или недостаточности тиамина развивается тяжелое заболевание – бери-бери, широко распространенное в ряде стран Азии и Индокитая, где основным продуктом питания является рис. Следует отметить, что недостаточность витамина B1встречается и в европейских странах, где она известна как симптом Вернике, проявляющийся в виде энцефалопатии, или синдром Вейса с преимущественным поражением сердечно-сосудистой системы. Специфические симптомы связаны с преимущественными нарушениями деятельности и сердечно-сосудистой, и нервной систем, а также пищеварительного тракта. В настоящее время пересматривается точка зрения, что бери-бери у человека является следствием недостаточности только витамина В1. Более вероятно, что это заболевание представляет собой комбинированный авитаминоз или полиавитаминоз, при котором организмиспытывает недостаток также в рибофлавине, пиридоксине, витаминах РР, С и др. На животных и добровольцах получен экспериментальный авитаминоз Bl. В зависимости от преобладания тех или иных симптомов различают ряд клинических типов недостаточности, в частности полиневритную (сухую) форму бери-бери, при которой на первый план выступают нарушения в периферической нервной системе. При так называемой отечной форме бери-бери преимущественно поражается сердечно-сосудистая система, хотя отмечаются также явления полиневрита. Наконец, выделяют остро протекающую кардиальную форму болезни, называемую пернициозной, которая приводит к летальному исходу в результате развития острой сердечной недостаточности. В связи с внедрением в медицинскую практику кристаллического препарата тиамина летальность резко сократилась и наметились рациональные пути лечения и профилактики этого заболевания. К наиболее ранним симптомам авитаминоза В1 относятся нарушения моторной и секреторной функций пищеварительного тракта: потеря аппетита, замедление перистальтики (атония) кишечника, а также изменения психики, заключающиеся в потере памяти на недавние события, склонности к галлюцинациям; отмечаются изменения деятельности сердечно-сосудистой системы: одышка, сердцебиение, боли в области сердца. При дальнейшем развитии авитаминоза выявляются симптомы поражения периферической нервной системы (дегенеративные изменения нервных окончаний и проводящих пучков), выражающиеся в расстройстве чувствительности, ощущении покалывания, онемения и болей по ходу нервов. Эти поражения завершаются контрактурами, атрофией и параличами нижних, а затем и верхних конечностей. В этот же период развиваются явления сердечной недостаточности (учащение ритма, сверлящие боли в области сердца). Биохимические нарушения при авитаминозе В1 проявляются развитием отрицательного азотистого баланса, выделением в повышенных количествах с мочой аминокислот и креатина, накоплением в крови и тканях α-кетокислот, а также пентозосахаров. Содержание тиамина и ТПФ в сердечной мышце и печени у больных бери-бери в 5-6 раз ниже нормы. Биологическая роль. Экспериментально доказано, что витамин B1в форме ТПФ является составной часть минимум 5 ферментов, участвующих в промежуточном обмене веществ. ТПФ входит в состав двух сложных ферментных систем – пируват - и α - кетоглутарат дегидрогеназных комплексов, катализирующих окислительноедекарбоксилирование пировиноградной и α-кетоглутаровой кислот. В составе транскетолазы ТПФ участвует в переносе гликоальдегидного радикала от кетосахаров на альдосахара (см. главу 10). ТПФ является коферментомпируватдекар-боксилазы клеток дрожжей (при алкогольной ферментации) и дегидро-геназы γ-оксикетоглутаровойкислоты. Приведенными примерами, вероятнее всего, не ограничиваются биологические функции тиамина. В частности, ТПФ участвует в окислительном декарбоксилировании глиоксиловой кислоты и α-кетокислот, образующихся при распадеаминокислот с разветвленной боковой цепью; в растениях ТПФ является эссенциальным кофактором при синтезевалина и лейцина в составе фермента ацетолактатсинтетазы. Распространение в природе и суточная потребность. Витамин В1 широко распространен в природе. Основное количество его человек получает с растительной пищей. Много витамина B1содержится в дрожжах, пшеничном хлебе из муки грубого помола, оболочке и зародышах семян хлебных злаков, сое, фасоли, горохе, меньше – в картофеле, моркови, капусте. Из продуктов животного происхождения наиболее богаты витамином B1печень, почки, мозг. Некоторые бактерии, населяющие кишечник животных, способны синтезировать достаточное количество тиамина: например, количества витамина В1, синтезированного микрофлорой кишечника коров, оказывается вполне достаточно для покрытия потребностей организма. Рекомендуемые Институтом питания РАМН нормы суточного потребления тиамина для отдельных групп населения составляют от 1,2 до 2,2 мг. 18. ИНГИБИРОВАНИЕ ФЕРМЕНТНОЙ АКТИВНОСТИ. Можно выделить два основных направления ингибирования · по прочности связывания фермента с ингибитором ингибирование бывает обратимым и необратимым. · по отношению ингибитора к активному центру фермента ингибирование делят на конкурентное и неконкурентное. Необратимое ингибирование При необратимом ингибировании происходит связывание или разрушение функциональных групп фермента, необходимых для проявления его активности. Например, вещество диизопропилфторфосфат прочно и необратимо связывается с гидроксигруппой серина в активном центре фермента ацетилхолинэстеразы, гидролизующей ацетилхолин в нервных синапсах. Ингибирование этого фермента предотвращает распад ацетилхолина в синаптической щели, в результате чего медиатор продолжает оказывать воздействие на свои рецепторы, что бесконтрольно усиливает холинергическую регуляцию. Аналогичным образом действуют боевые фосфоорганические вещества (зарин, зоман) и инсектициды (карбофос, дихлофос).
Механизм необратимого ингибирования ацетилхолинэстеразы Еще один пример связан с ингибированием ацетилсалициловой кислотой (аспирином) ключевого фермента синтеза простагландинов – циклооксигеназы. Эта кислота входит в состав противовоспалительных средств и используется при воспалительных заболеваниях и лихорадочных состояниях. Присоединение ацетильной группы к аминогруппе в активном центре фермента вызывает инактивацию последнего и прекращение синтеза простагландинов.
Механизм необратимого ингибирования циклооксигеназы Обратимое ингибирование При обратимом ингибировании происходит непрочное связывание ингибитора с функциональными группами фермента, вследствие чего активность фермента постепенно восстанавливается. Примером обратимого ингибитора может служить прозерин, связывающийся с ферментом ацетилхолинэстеразой в ее активном центре. Группа ингибиторов холинэстеразы (прозерин, дистигмин, галантамин) используется при миастении, после энцефалита, менингита, травм ЦНС. Конкурентное ингибирование При таком виде ингибирования ингибитор по своей структуре похож на субстрат фермента. Поэтому он соперничает с субстратом за активный центр, что приводит к уменьшению связывания субстрата с ферментом и нарушению катализа. В этом состоит особенность конкурентного ингибирования – возможность усилить или ослабить ингибирование через изменение концентрации субстрата. Например: 1. Конкурентное взаимодействие этанола и метанола за активный центр алкогольдегидрогеназы. 2. Ингибирование сукцинатдегидрогеназы малоновой кислотой, структура которой схожа со структурой субстрата этого фермента – янтарной кислоты (сукцината).
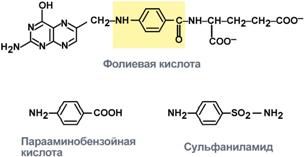
Конкурентное ингибирование сукцинатдегидрогеназы 3. Также к конкурентным ингибиторам относят антиметаболиты или псевдосубстраты, например, антибактериальные средства сульфаниламиды, схожие по структуре с п -аминобензойной кислотой, компонентом фолиевой кислоты. При лечении сульфаниламидами в бактериальной клетке конкурентно нарушается использование п -аминобензойной кислоты для синтеза фолиевой кислоты, что и вызывает лечебный эффект.
Сходство строения сульфаниламидов ОБРАЗОВАНИЕ ЖЕЛЧНЫХ КИСЛОТ. БИОРОЛЬ. Источником образования желчных кислот является холестерин. Печень синтезирует холевую и хенодезоксихолевую кислоты, однако в желчи встречаются и дезоксихолевая, липохолевая, а также урсодезоксихолевая кислоты, которые образуются в кишечнике при воздействии бактериальной флоры из первичных желчных кислот. Благодаря особому строению (наличие гидрофильных и гидрофобных компонентов в молекуле) желчные кислоты выступают стабилизаторами физико-химического состояния желчи. Способность их образовывать мицеллы и растворять липидные соединения (в том числе холестерин) играет значительную роль в переваривании и абсорбции пищевых жиров. Эти свойства желчных кислот используются в терапии желчно-каменной болезни в целях растворения холестериновых камней. Секреция компонентов желчи мембраной гепатоцита протекает при активном транспорте органических веществ и электролитов и пассивном переносе молекул воды. Синтез желчных кислот Образование желчных кислот идет в эндоплазматическом ретикулуме при участии цитохрома Р450, кислорода, НАДФН и аскорбиновой кислоты. 75% холестерина, образуемого в печени, участвует в синтезе желчных кислот.
Реакции синтеза желчных кислот на примере холевой кислоты В печени синтезируются первичные желчные кислоты – холевая (гидроксилирована по С3, С7, С12) и хенодезоксихолевая (гидроксилирована по С3, С7), затем они образуют конъюгаты с глицином – гликопроизводные и с таурином – тауропроизводные, в соотношении 3:1 соответственно.
Строение желчных кислот В кишечнике под действием микрофлоры эти желчные кислоты теряют НО-группу при С7 и превращаются во вторичные желчные кислоты – дезоксихолевую (гидроксилирована по С3 и С12) и литохолевую (гидроксилирована только по С3). ВИТАМИН В6 Витамин В6 был открыт П.Дьерди в 1934 г., а через четыре года выделен в кристаллическом состоянии. Различают три индивидуальных вещества. Обладающих свойствами Витамин В6: пиридоксамин, пиридоксин и пиридоксаль. Кристаллы пиридоксамина бесцветные с температурой плавления 160 0С, хорошо растворимы в воде и некоторых органических растворителях. Аналогичными свойствами обладает и пиридоксаль. Кристаллы пиридоксина плавятся при температуре 195 0С и почти не растворяются в органических растворителях.
1.1 Активные формы. Все три формы витамина В6 легко превращаются друг в друга, однако наибольшую биологическую значимость имеет фосфорилированная форма пиридоксаля. Витамин В6 синтезируется растительными и микробными клетками. Наибольшее количество его в печени, яйцах, дрожжах, моркови. Суточная потребность человека в этом витамине 3-4 мг. Из антивитаминов В6 наибольшее значение имеет изониазид, так как он оказывает антибактериальное действие на туберкулёзные микобактерии и используется в качестве противотуберкулёзного препарата.
1.2 Метаболизм. После поступления с пищей в организм витамин всасывается в тонком кишечнике методом простой диффузии. С током крови он транспортируется к тканям и достаточно легко проникает вовнутрь клеток. В клетках происходит фосфорилирование всех трех витамина В6 , однако наибольшую значимость имеет фосфопиридоксаль. Фосфорилирование осуществляется при помощи фермента пиридоксалькиназы, при этом образуется активная форма витамина В6 или кофермент фосфопиридоксаль. Катаболизм фосфопиридоксаля начинается с фосфорилирования, которое катализируется различными фосфотазами. Далее происходит окисление и образуется 4-пиридоксиловая кислота, которая выводится из организма с мочой. 1.3 Биохимические функции. Основная функция витамина В6 состоит в том, что он в составе различных ферментов принимает участие в метаболизме аминокислот. Его активная форма – фосфопиридоксаль, является коферментом многих ферментов аминокислотного обмена. Фосфопиридоксаль входит в состав аминотрансфераз, катализирующих перенос аминогрупп ряда аминокислот. Кроме того идентифицировано более 50 пиридоксальфосфатных ферментов, катализирующих разнообразные реакции, такие, как декарбоксилирование аминокислот, рацемизация, дегидротирование, гидролитическое расщепление субстратов. Фосфопиридоксалевые ферменты катализируют синтез сфингозина, (это липидный обмен). Витамин В6 влияет на активность ряда ферментов углеводного обмена, в частности его дефицит является причиной снижения активности глюкозо-6-фосфатдегидрогеназы. Витамин В6 активирует работу гликогенфосфорелазы (распад гликогена). Кроме того, установлено влияние витамина В6 на метаболизм жирных кислот. Витамин В6 катализирует реакции: А) трансаминирования (самая активная АЛТ, АСТ). Б)Декарбоксилирование синтез биогенных аминов. В) Витамин В6 принимает участие в обмене серосодержащих аминокислот. Витамин В6 совместно с витамином В2 принимают участие в обмене триптофана,следствие чего синтез НАД+, в состав которого входит витамин РР (никотинамид). 1.4 Гипо- и гипервитаминоз. Гиповитаминоз. Признаком гиповитаминоза являются кожные заболевания (эритемы, эдемы), нарушения центральной нервной системы и кроветворения. Дефицит витамина В6 у человека проявляется более отчетливо в младенческом возрасте и сопровождается конвульсиями и эпилептическими припадками. Гипервитаминоз. В большинстве случаев прием дозы, даже в 10 раз превышающей потребность в витамине В6 (2 мг/сут), безопасен. Если в течение длительного времени принимать несколько граммов витамина В6 в сутки, развиваются тяжелые нейропатии, проявляющиеся атаксией, онемением губ, неловкостью движений рук, неловкостью движений ног, нарушениями проприоцептивной чувствительности, нарушениями вибрационной чувствительности (другие виды чувствительности и рефлексы не изменены). 1.5 Практическое применение. В медицинской практике используют как витамин В6 , так и его коферментные формы. Пиридоксин применяют при токсикозах у беременных, атеросклерозе, нервных и кожных заболеваниях. Пиридоксальфосфат более эффективен, особенно при кожных заболеваниях. ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ Цикл трикарбоновых кислот (цикл лимонной кислоты, цикл Кребса). Это циклическая последовательность ферментативных реакций, в которой участвуют ди- и трикарбоновые кислоты, являющиеся промежуточными продуктами при распаде белков, жиров и углеводов у животных, растений и микробов. Таким образом, цикл трикарбоновых кислот – конечный (терминальный) общий путь окисления белков, липидов и углеводов. Он происходит в митохондриях. Открыт в 1937 г. Х. Кребсом и У. Джонсоном. Цикл трикарбоновых кислот поставляет вещества для различных биосинтезов. Он осуществляется в аэробных условиях. Лимитирующей стадией цикла является синтез янтарной кислоты (сукцината). Первая стадия цикла представляет собой альдольную конденсацию щавелевоуксусной (кетоянтарной) кислоты с ацетилкоферментом А и последующий гидролиз тиоэфирной связи С~S. Суммарное уравнение цикла трикарбоновых кислот: СН3СО~SСоА + 3НАД+ + ФАД + ГДФ + Н3РО4 + 2Н2О ® 2СО2 + 3НАДН + 2Н+ + ФАДН2 + ГТФ + НSСоА. Ё ОБМЕН МЕТИОНИНА. ОБРАЗОВАНИЕ SAM (В ТЕТРАДИ). ПРИМЕНЕНИЕ В МЕДИЦИНЕ. Реакция активация метионина Активной формой метионина является S-аденозилметионин (SAM) - сульфониевая форма аминокислоты, образующаяся в результате присоединения метионина к молекуле аденозина. Аденозин образуется при гидролизе АТФ (см. схему А). Эту реакцию катализирует фермент метионин аденозилтрансфераза, присутствующий во всех типах клеток. Структура (-S+-CH3) в SAM - нестабильная группировка, определяющая высокую активность метильной группы (отсюда термин "активный метионин"). Эта реакция уникальна для биологических систем, так как, по-видимому, является единственной известной реакцией, в результате которой освобождаются все три фосфатных остатка АТФ. Отщепление метильной группы от SAM и перенос её на соединение-акцептор катализируют ферменты метилтрансферазы. SAM в ходе реакции превращается в S-аденозилгомоцистеин (SAT).
СИНТЕЗ ГЛИКОГЕНА. Прежде всего глюкоза подвергается фосфорилированию при участии фермента гексокиназы, а в печени – и глюкокиназы. Далее глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазы переходит в глюкозо-1-фос-фат:
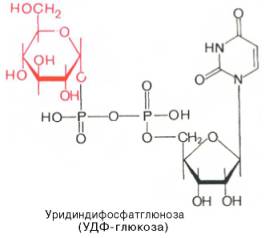
Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в синтез гликогена. На первой стадии синтеза глюкозо-1-фосфат вступает во взаимодействие с УТФ (уридинтрифосфат), образуя уридиндифосфатглюкозу (УДФ-глюкоза) и пирофосфат. Данная реакция катализируется ферментом глюкозо-1-фосфат-уридилилтрансферазой (УДФГ-пирофосфорилаза): Глюкозо-1-фосфат + УТФ < = > УДФ-глюкоза +Пирофосфат. Приводим структурную формулу УДФ-глюкозы:
На второй стадии – стадии образования гликогена – происходит перенос глюкозного остатка, входящего в состав УДФ-глюкозы, на глюкозидную цепь гликогена («затравочное» количество). При этом образуется α-(1–>4)-связь между первым атомом углерода добавляемого остатка глюкозы и 4-гидроксильной группой остатка глюкозы цепи. Этареакция катализируется ферментом гликогенсинтазой. Необходимо еще раз подчеркнуть, что реакция, катализируемая гликогенсинтазой, возможна только при условии, что полисахаридная цепь уже содержит более 4 остатков D-глю-козы.
Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и таким образом весь цикл превращений глюкозо-1-фосфата начинается сначала. В целом образование α-1,4-глюкозидной ветви («амилозной» ветви) гликогена можно представить в виде следующей схемы:
Установлено, что гликогенсинтаза неспособна катализировать образование α-(1–>6)-связи, имеющейся в точках ветвления гликогена. Этот процесс катализирует специальный фермент, получивший название гли-когенветвящегофермента, или амило-(1–>4)–>(1–>6)-трансглюкозидазы. Последний катализирует перенос концевого олигосахаридного фрагмента, состоящего из 6 или 7 остатков глюкозы, с нередуцирующего конца одной из боковых цепей, насчитывающей не менее 11 остатков, на 6-гидроксиль-ную группу остатка глюкозы той же или другой цепигликогена. В результате образуется новая боковая цепь. Ветвление повышает растворимость гликогена. Кроме того, благодаря ветвлению создается большое количество невосстанавливающих концевых остатков, которые являются местами действия гликогенфосфорилазы и гликогенсинтазы. Таким образом, ветвление увеличивает скорость синтеза и расщепления гликогена. Благодаря способности к отложению гликогена (главным образом в печени и мышцах и в меньшей степени в других органах и тканях) создаются условия для накопления в норме некоторого резерва углеводов. При повышении энерготрат в организме в результате возбуждения ЦНС обычно происходят усиление распада гликогена и образование глюкозы. Помимо непосредственной передачи нервных импульсов к эффекторным органам и тканям, при возбуждении ЦНС повышаются функции ряда желез внутренней секреции (мозговое вещество надпочечников, щитовидная железа,гипофиз и др.), гормоны которых активируют распад гликогена, прежде всего в печени и мышцах (см. главу 8). Как отмечалось, эффект катехоламинов в значительной мере опосредован действием цАМФ, который активируетпротеинкиназы тканей. При участии последних происходит фосфорилирование ряда белков, в том числе гликогенсинтазы и фосфорилазы b – ферментов, участвующих в обмене углеводов. Фосфорилированный ферментгликогенсинтаза сам по себе малоактивен или полностью неактивен, но в значительной мере активируется положительным модулятором глюкозо-6-фосфатом, который увеличивает Vmaxфермента. Эта форма гликогенсинтазы называется D-формой, или зависимой (dependent) формой, поскольку ее активность зависит от глюкозо-6-фосфата. Дефосфорилированная форма гликоген-синтазы, называемая также I-формой, или независимой (independent) формой, активна и в отсутствие глюкозо-6-фосфата. Таким образом, адреналин оказывает двойное действие на обмен углеводов: ингибирует синтез гликогена из УДФ-глюкозы, поскольку для проявления максимальной активности D-формы гликогенсинтазы нужны очень высокиеконцентрации глюкозо-6-фосфата, и ускоряет распад гликогена, так как способствует образованию активной фосфорилазы а. В целом суммарный результат действия адреналина состоит в ускорении превращения гликогена вглюкозу. ПЕРЕВАРИВАНИЕ УГЛЕВОДОВ.
|
|||||||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 1467; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.129.210.17 (0.1 с.) |