Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Поражения больших полушарий и расстройства высших корковых функций
В большом мозге человека мы различаем рассмотренные уже подкорковые ганглии основания, белое вещество полушарий и, наконец, кору большого мозга, представляющую собой наиболее поздний по развитию и наиболее совершенный отдел центральной нервной системы. Анатомически кора представляет собой пластину серого вещества, покрывающую наружную поверхность полушарий. Различают наружную (выпуклую) поверхность полушарий, внутреннюю их поверхность и основание. На наружной поверхности мощная роландова борозда (рис. 88) разделяет лобную и теменную доли. Ниже ее расположенная сильвиева борозда отделяет височную долю от теменной и лобной. Затылочная доля отделяется от теменной и височной линией, продолжающейся книзу от sulcus parietooccipitalis. Таким образом, на выпуклой поверхности каждого полушария намечаются четыре доли коры головного мозга: лобная, теменная, височная и затылочная (по другому делению — еще лимбическая и островок). Две массивные извилины, расположенные «по берегам» роландовой борозды, одна — кпереди от последней (передняя центральная извилина) и другая - кзади от нее (задняя центральная извилина), часто выделяются в особую долю, именуемую областью центральных извилин. На наружной, выпуклой поверхности различают: в собственно лобной доле (в области кпереди от передней центральной извилины) три извилины, расположенные примерно горизонтально: первую, или верхнюю, вторую, или среднюю, и третью, или нижнюю, лобные извилины (см. рис. 88). Теменная доля идущей в середине ее горизонтальной бороздой (sulcus interparietalis) делится на верхнюю и нижнюю теменные дольки. В нижней теменной дольке различают расположенную более кпереди gyrus supramarginalis и кзади от нее граничащую с затылочной долей gyrus angularis. В височной доле заметны три горизонтально расположенные извилины: первая, или верхняя, вторая, или средняя, и третья, или нижняя, височные извилины. На внутренней поверхности полушарий после разреза мозга по сагиттальной линии (рис. 89) хорошо выражена sulcus parietooccipitalis, отделяющая затылочную долю от теменной. В затылочной доле намечена глубокая fissura calcarina (sulcus calcarinus — PNA), выше которой
Рис. 88. Наружная поверхность полушария головного мозга. расположен cuneus и ниже gyms lingualis seu occipitotemporalis medialis (PNA). В переднем отделе височной доли находится gyrus hyppocampi (parahippocampalis — PNA). В середине среза видны пересеченные волокна основной комиссуральной спайки полушарий — corporis callosi (мозолистого тела).
На нижней поверхности полушарий большого мозга (на основании) в переднем отделе находятся лобные доли, кзади от них — отделенные сильвиевой бороздой височные и еще более кзади — затылочные доли (см. рис. 90). На основании лобных долей видны bulbus и tractus olfac-torii, кзади от них — перекрест зрительных нервов (chiasma opticum). Достаточно массивным образованием является мозговой ствол: ножки мозга, варолиев мост, продолговатый мозг и лежащий над последними и под затылочными долями мозжечок. Кора представляет собой серое вещество. Микроскопическое строение ее довольно сложное; кора состоит из ряда слоев клеток и их волокон.
Рис. 89. Внутренняя поверхность полушария головного мозга.
Рис. 90. Основание головного мозга. Основной тип строения мозговой коры — шестислойный (рис. 91). I. Молекулярный слой, самый поверхностный, лежит непосредственно под мягкой мозговой оболочкой, беден клетками, волокна его имеют параллельное поверхности коры направление, отчего он носит еще наименование тангенциального. II. Наружный зернистый слой расположен глубже первого, включает большое количество мелких зернистых нервных клеток. III. Слой малых и средних пирамидных клеток. IV. Внутренний зернистый слой. V. Слой больших пирамидных клеток. VI. Слой полиморфных клеток состоит из клеток самой разнообразной формы (треугольных, веретенообразных и др.). Как видно на рисунке 91, волокна перечисленных клеток имеют либо параллельное поверхности коры направление (ассоциационные пути, связывающие между собой различные территории коры), либо являются радиальными, перпендикулярными к поверхности. Последнего типа волокна типичны для проекционных путей (связывающих кору головного мозга с лежащими ниже ее образованиями). Шестислойный тип строения коры является далеко не однородным. Существуют участки коры, в которых один из слоев представляется особенно мощным, а другой — весьма слабо выраженным. В других областях коры намечается подразделение некоторых слоев на подслои.
Основоположником детального изучения строения клеточного состава коры был русский ученый В.А.Бец. За много лет до Бродманна, Фохта, Экономо и др. с помощью нового, разработанного им самим метода серийных срезов мозга и окраски кармином В.А.Бецем была тщательно разработана цитоархитектоника коры головного мозга и сделан большой шаг в учении о локализации функции в ней. Сам термин «архитектоника» коры принадлежит также В.А.Бецу'. 1 Интересно отметить, что в своем докладе в 1949 г. в Париже американский ученый Бейли признал, что последние тщательные его исследования цитоархитектоники коры показали полное совпадение со старыми описаниями В.А.Беца, «атлас, которых, к несчастью, никогда напечатан не был». Бейли даже высказывает сомнение, что последующие работы дали реальный прогресс в этом направлении (после указанных описаний В.А.Беца и Флексига).
Рис. 91. Цитоархитектоническая и миелоархитектоническая схема коры головного мозга. Весьма многочисленные последующие работы, особенно отечественных ученых (И.Н.Филимонов, С.А.Саркисов, Е.Т.Кононова и др.), расширили наши представления о тонкой гистологической структуре коры головного мозга. На рисунках 92 и 93 изображены цитоархитек-тонические поля коры человека (Институт мозга, Москва). Результаты цитоархитектонических исследований сыграли известную роль в решении спорных вопросов о локализации функций в коре больших полушарий. Установлено, что области, связанные с определенной функцией, имеют свое, свойственное им строение; что участки коры, близкие по своему функциональному значению, имеют известное сходство в строении как у животных, так и у человека. Те же участки, поражения которых вызывают расстройство сложных, чисто человеческих функций (например, речевых), имеются только в коре человека, а у млекопитающих, в том числе даже у антропоморфных обезьян, отсутствуют. ЛОКАЛИЗАЦИЯ ФУНКЦИЙ В КОРЕ Представление о локализации функций в коре головного мозга имеет большое практическое значение для решения задач топики поражений в больших полушариях мозга. Однако до сего времени многое в этом разделе остается еще спорным и не вполне разрешенным. Учение о локализации функций в коре имеет довольно большую историю - от
Рис. 92. Цитоархитектоническая карта коры головного мозга человека. Наружная поверхность (Московский институт мозга).
Рис. 93. Цитоархитектоническая карта коры мозга человека. Внутренняя по- | верхность (Московский институт мозга). f отрицания локализованное™ в ней функций до распределения в коре на строго ограниченных территориях всех функций человеческой деятельности вплоть до самых высших качеств последней (памяти, воли и т.д.) и, наконец, до возвращения к «эквипотенциальное™» коры, т.е. снова, по существу, к отрицанию локализации функций (в последнее время за рубежом). Представления о равнозначности (эквипотенциальное™) различных корковых полей вступают в противоречие с огромным фактическим материалом, накопленным морфологами, физиологами и клиницистами. Повседневный клинический опыт показывает, что существуют определенные незыблемые закономерные зависимости расстройств функций от расположения патологического очага. Исходя из этих основных положений, клиницист и решает задачи топической диагностики. Однако так обстоит дело до тех пор, пока мы оперируем с расстройствами, относящимися к сравнительно простым функциям: движениям, чувствительности и др. Иначе говоря, твердо установленной является локализация в проекционных зонах — корковых полях, непосредственно связанных своими путями с нижележащими отделами нервной системы и периферией. Функции коры более сложные, филогенетически более молодые, не могут быть узко локализованными; в осуществлении сложных функций участвуют весьма обширные области коры, и даже вся кора в целом. Вот почему решение задач топики поражений на основании расстройств речи, апраксии, агнозии и тем более психических нарушений, как показывает клинический опыт, более затруднительно и иногда неточно.
Вместе с тем в пределах мозговой коры имеются участки, поражение которых вызывает тот или иной характер, ту или иную степень, например, речевых расстройств, нарушений гнозии и праксии, топодиа-гностическое значение которых также является значительным. Из этого, однако, не следует, что существуют специальные, узко локализованные центры, «управляющие» этими сложнейшими формами человеческой деятельности. Необходимо четко разграничивать локализацию функций и локализацию симптомов. Основы нового и прогрессивного учения о локализации функций в головном мозге были созданы И.П.Павловым. Вместо представления о коре больших полушарий как в известной мере изолированной надстройке над другими этажами нервной системы с узко локализованными, связанными по поверхности (ассоциаци-онными) и с периферией (проекционными) областями И.П.Павлов создал учение о функциональном единстве нейронов, относящихся к различным отделам нервной системы, - от рецепторов на периферии до коры головного мозга — учение об анализаторах. То, что мы называем центром, является высшим, корковым отделом анализатора. Каждый анализатор связан с определенными областями коры головного мозга (рис. 94, см. вклейку). И.П.Павлов вносит существенные коррективы в прежние представления об ограниченности территорий корковых центров, в учение об узкой локализации функций. Корковое представительство анализаторов не ограничивается зоной проекций соответствующих проводников, а распространяется на более широкие области; корковые поля различных анализаторов перекрывают друг друга (см. рис. 94).
С этим выводом, основанным на обширных экспериментально-физиологических исследованиях, вполне согласуются новейшие морфологические данные о невозможности точного разграничения корковых цитоархитектонических полей. Следовательно, функции анализаторов (или, иными словами, работу первой сигнальной системы) нельзя связывать только с корковыми проекционными зонами (ядрами анализаторов). Тем более нельзя узко локализовать сложнейшие, чисто человеческие функции — функции второй сигнальной системы. Работа второй сигнальной системы неразрывно связана с функциями всех анализаторов, поэтому невозможно представить локализацию сложных функций второй сигнальной системы в каких-либо ограниченных корковых полях. Значение наследства, оставленного нам великим физиологом, для правильного развития учения о локализации функций в коре головного мозга исключительно велико. И.П.Павловым заложены основы учения о динамической локализации функций в коре. Представления о динамической локализации предполагают возможность использования одних и тех же корковых структур в разнообразных сочетаниях для обслуживания различных сложных корковых функций. Сохраняя ряд упрочившихся в клинике определений и истолкований, мы попытаемся внести в наше изложение некоторые коррективы в свете учения И.П.Павлова о нервной системе и ее патологии. Прежде всего, нужно рассмотреть вопрос о так называемых проекционных и ассоциационных центрах. Привычное представление о двигательных, чувствительных и других проекционных центрах (передней и задней центральных извилинах, зрительных, слуховых центрах и др.) связано с понятием о довольно ограниченной локализации в данной области коры той или иной функции, причем этот центр непосредственно связан с нижележащими нервными приборами, а в последующем и с периферией, своими проводниками (отсюда и определение — «проекционный»). Примером такого центра и его проводника является, например, передняя центральная извилина и пирамидный путь; sulcus calcarinus, radiatio oplica и т.д. Проекционные центры ассо-циационными путями связаны с другими центрами, с поверхностью коры. Эти широкие и мощные ассоциационные пути и обусловливают возможность сочетанной деятельности различных корковых областей, установления новых связей, формирования, следовательно, условных рефлексов.
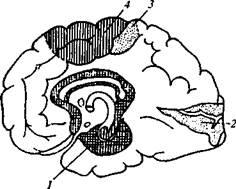
Рис, 95. Выпадения функций, наблюдаемые при поражении различных отделов коры головного мозга (наружной поверхности). 2 - расстройства зрения (гемианоп-сия); 3 — расстройства чувствительности; 4 — центральные параличи или парезы; 5- аграфия; 6- корковый паралич взгляда и поворота головы в противоположную сторону; 7— моторная афазия; 8 - расстройства слуха (при одностороннем поражении не наблюдаются); 9 - ам-нестическая афазия; 10 — алексия; // - зрительная агнозия; 12 - асте-реогнозия; 13 — апраксия; 14— сенсорная афазия.
«Ассоциационные центры», в отличие от проекционных, непосредственной связи с нижележащими отделами нервной системы и периферией не имеют; они связаны только с другими участками коры, в том числе и с «проекционными центрами». Примером «ассо-циационного центра» может служить так называемый «центр стерео-гнозии» в теменной доле, расположенный кзади от задней центральной извилины (рис. 95). Деление центров на проекционные и ассоциационные представляется нам неправильным. Большие полушария представляют собой совокупность анализаторов. Верхние слои коры больших полушарий, по сути говоря, целиком заняты воспринимающими центрами или, по терминологии И.П.Павлова, «мозговыми концами анализаторов»1. Вместе с тем от всех долей, от нижних слоев коры идут уже эфферентные проводники, соединяющие корковые концы анализаторов с исполнительными органами через посредство подкорковых, стволовых и спинальных аппаратов. Примером такого афферентного проводника является пирамидный путь — этот вставочный неврон между кинестетическим (двигательным) анализатором и периферическим двигательным невроном. Передняя центральная извилина является лишь двигательной проекционной областью для пирамидных путей, а не «проекционным двигательным центром». Не подлежит сомнению, что «произвольные» движения есть условные двигательные рефлексы, т.е. движения, сложившиеся, «проторенные», в процессе индивидуального жизненного опыта; но в выработке, организации и уже создавшейся деятельности скелетной мускулатуры все зависит от афферентного прибора - кожного и двигательного анали- 1 С этим следует сопоставить тог факт, что афферентные проводниковые системы количественно 1 5-6 раз превышают эфферентные. затора (клинически — кожной и суставно-мышечной чувствительности, шире — кинестетического чувства), без которого невозможна тонкая и точная координация двигательного акта. Двигательный анализатор (задача которого - анализ и синтез «произвольных» движений) совершенно не соответствует представлениям о корковых двигательных «проекционных» центрах с определенными границами последних и четким соматотопическим распределением. Двигательный анализатор, как и все анализаторы, связан с очень широкими территориями коры, и двигательная функция (в отношении «произвольных» движений) чрезвычайно сложна. Эта сложная функция отнюдь не обеспечивается только проекционной зоной для пирамидных путей. Так же обстоит дело и с другими «проекционными» центрами (кожной чувствительности, зрения, слуха, вкуса, обоняния). Следовательно, в каждом анализаторе (корковом его отделе), в том числе и двигательном, существует область или зона, «проецирующая» на периферию (двигательная область) или в которую «проецируется» периферия (чувствительные области и в том числе кинестетические рецепторы для двигательного анализатора). Допустимо, что «проекционное ядро анализатора» можно отождествить с понятием о двигательной или чувствительной проекционной зоне. Максимум нарушений, писал И.П.Павлов, анализа в синтезе возникает при поражении именно такого «проекционного ядра»; если принять за реальную максимальную «поломку» анализатора максимум нарушения функции, что объективно является совершенно правильным, то наибольшим проявлением поражения двигательного анализатора является центральный паралич, а чувствительного — анестезия. С этой точки зрения правильным будет понятие «ядро анализатора» отождествить с понятием «проекционная область анализатора». На основании изложенного считаем правильным заменить понятие о проекционном центре понятием о проекционной области в зоне анализатора. Тогда деление корковых «центров» на проекционные и ассо-циациоиные необоснованно: существуют анализаторы (корковые и их отделы) и в их пределах — проекционные области. Проекционные области коры Двигательные проекционные области для мускулатуры противоположной стороны тела расположены в передней центральной извилине (см. рис. 94 и 95, 4). Проекция для отдельных мышечных групп представлена здесь в порядке, обратном расположению их в теле: верхним отделам передней центральной извилины (и частично lobulus paracentralis на внутренней поверхности полушария - рис. 96) соответствует нога, средним отделам - рука и нижним отделам передней центральной извилины - лицо,
Рис. 96. Выпадения функций, наблюдаемые при поражении различных отделов коры (внутренней поверхности). / - расстройства обоняния (при одностороннем поражении не наблюдаются); 2 — расстройства зрения (гемианоп-сии); 3 - расстройства чувствительности; 4- центральные параличи или парезы. язык, гортань и глотка. Проекция движений туловища, по-видимому, представлена в заднем отделе верхней лобной извилины1. Проекции поворота глаз и головы в противоположную сторону соответствует задний отдел второй лобной извилины. Пути отсюда направляются книзу в непосредственной близости к пирамидным пучкам и устанавливают связи с задним продольным пучком в мозговом стволе, осуществляя произвольную иннервацию взгляда (одного его или в сочетании с поворотом головы)2. Чувствительные проекционные области находятся в задней центральной извилине (см. рис. 94 и 95, J). Проекция кожных рецепторов аналогична соматотопическому представительству в передней центральной извилине: в верхнем отделе извилины представлена чувствительность нижней конечности, в среднем - руки и в нижнем - головы. Зрительная проекционная область расположена в затылочных долях, на внутренней поверхности полушарий, по краям и в глубине sulci calcari-ni (см. рис. 96, 2). В каждом полушарий представлены противоположные поля зрения обоих глаз, причем область, расположенная над sulco calcari-no (cuneus), соответствует нижним, а область под ней (gyms lingualis, или occipitotemporalis medialis — PNA) — верхним квадрантам полей зрения. Слуховая проекционная область коры находится в височных долях, в первой (верхней) височной извилине и в извилинах Гешля (на внутренней поверхности височной доли). Обонятельная проекционная область расположена также в височных долях, главным образом в gyms parahippocampalis, в особенности в его переднем отделе (крючке, или uncus) (см. рис. 96, 7). 1 Возможны варианты распределения корковой проекции для мускулатуры тела. Клинические данные и опыт электрического раздражения обнаженной коры при операциях нередко показывают расположение территорий для ноги полностью на внутренней поверхности полушарий; в этих случаях К'рритории для руки смещены кверху и доходят до верхнего края передних центральных извилин. - Другая добавочная область, раздражение которой вызывает поворот глаз и головы в противоположную сторону, находится на границе теменной и затылочной долей. Также в височной доле близко к обонятельным территориям расположены и вкусовые1. Все проекционные области коры являются двусторонними, симметрично расположенными в каждом полушарии. Часть из них связана только с противоположной стороной (передняя и задняя центральные извилины, зона поворота глаз и головы, зрительная область). Корковые слуховые, обонятельные и вкусовые территории каждого полушария связаны с соответствующими рецепторными полями на периферии с обеих сторон (противоположной и своей). Импульсы «произвольных» движений (результат корковой, услов-норефлекторной деятельности) через систему нисходящих проводников проходят к скелетной мускулатуре. От периферии — широкого поля рецепторов кожных, мышечных, специальных органов чувств, висцеральных и других — центростремительные импульсы через систему афферентных путей входят в соответствующие проекционные чувствительные территории, к специальным клеткам определенного слоя коры, и отсюда - в пределы широких территорий анализатора, где и происходит анализ и синтез восприятия. Правильным будет ограничить понятие проекционной области лишь группой специальных нервных клеток, расположенных на известной территории, только в определенном клеточном слое коры. Только площадью этой клеточной группы и определяется проекционная область. В остальном вся кора этой территории, несомненно, входит в состав данного анализатора. Таким образом, передняя, скажем центральная, извилина полностью входит в широкие поля двигательного анализатора, задняя центральная извилина — кожного и т.д.
|
|||||||||
|
|
Последнее изменение этой страницы: 2017-01-20; просмотров: 399; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.14.6.194 (0.042 с.) |