Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Занятие 31слуховая сенсорная системаСтр 1 из 4Следующая ⇒
ЗАНЯТИЕ 31СЛУХОВАЯ СЕНСОРНАЯ СИСТЕМА Особенности строения и свойств звуковоспринимающего и звукопроводящего аппаратов, обеспечивающих функцию слуха.
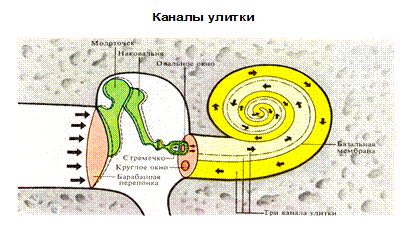
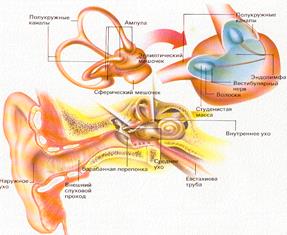
Периферическим отделом слухового анализатора, превращающим энергию звуковых волн в энергию нервного возбуждения, являются рецепторные волосковые клетки кортиева органа(орган Корти), находящегося в улитке. Слуховые рецепторы (фонорецепторы) относятся к механорецепторам, являются вторичными и представлены внутренними и наружными волосковыми клетками. Внутреннее(звуковоспринимающий аппарат), а также среднее(звукопередающий аппарат) и наружное ухо (звукоулавливающий аппарат) объединяются в понятие орган слуха. Т.о. орган слуха состоит из 3 отделов - наружного, среднего и внутреннего уха. Наружное и среднее ухо это вспомогательные сенсорные структуры, обеспечивающие проведения звука к слуховым рецепторам в улитке. Периферический отдел представлен ушной раковиной, наружным слуховым проходом и барабанной перепонкой. Среднее ухо (звукопроводящий отдел). Колебания барабанной перепонки передаются через заполненную воздухом полость среднего уха по цепи из 3-х маленьких слуховых косточек ( молоточка, наковальни и стремечка ) на овальное окно. Рукоятка молоточка вплетена в барабанную перепонку, а стремечко прикрепляется к мембране овального окна. Площадь барабанной перепонки (70 мм2), значительно больше площади овального окна(3,2 мм2), благодаря чему происходит усиление давления звуковых волн на мембрану овального окна примерно в 25 раз. В стенке, отделяющей среднее ухо от внутреннего кроме овального, есть еще круглое окно, тоже закрытое мембраной. Колебания жидкости улитки возникающее у овального окна, прошедшее по ходам улитки достигает, не затухая круглого окна. Мышцы среднего уха. Функциональная роль мышц среднего уха заключается в уменьшении воздействия громких звуков на слуховую систему. В среднем ухе имеются специальный защитный механизм, который представлен двумя мышцами – мышцей напрягающей барабанную перепонку (тимпанальная), т.е. ограничивают ее колебание при сильных звуках и мышцей фиксирующей стремечко (стременная), которая фиксирует стремечко и тем самым ограничивает его движения. Звукопонижающий рефлекс может уменьшать интенсивность передачи низкочастотных звуков на 30–40дБ, в то же самое время не затрагивая восприятия громкого голоса и шепотной речи. Значение этого рефлекторного механизма двояко: защита улитки от повреждающего вибрационного действия низкого звука и маскировка низких звуков в окружающей среде. Кроме того, мышцы слуховых косточек уменьшают чувствительность слуха человека к его собственной речи в момент, когда мозг активирует голосовой механизм.
Внутреннее ухо находится в височной кости. Представлено улиткой, в которой находится рецепторный аппарат. Она представляет собой костный спиральный, постепенно расширяющийся канал, образующий у человека 2,5 витка, который разделен основной мембраной и мембраной Рейснера на три узкие части (лестницы). Верхний канал (вестибулярная лестница) начинается от овального окна, соединяется с нижним каналом (барабанной лестницей) через геликотрему (отверстие в верхушке) и заканчивается круглым окном. Таким образом, верхние и нижние каналы представляют собой единый канал, начинающийся овальным окном и заканчивающийся круглым. Верхний и нижний канальцы заполнены перилимфой (похожа на спиномозговую жидкость по составу) в которой много Nа+ и мало К+. В средней лестнице или перепончатом канале в эндолимфе - в 100 раз больше К+и в 10 меньше Nа+, поэтому эндолимфа заряжена положительно по отношению к перилимфе. Это является причиной разности потенциалов.
Механизм слуховой рецепции Внутри среднего канала улитки на основной мембране расположен звуковоспринимающий аппарат - спиральный Кортиев орган. Основная мембрана вблизи овального окна по ширине составляет 0,04 мм, затем по направлению к вершине она постепенно расширяется, достигая у геликотреммы 0,5 мм. Здесь располагаются 2 вида рецепторных волосковых клеток: внутренние и наружные, отделенные друг от друга кортиевыми дугами. Внутренние волосковые клетки располагаются в один ряд (общее число их достигает 3500), наружные волосковые клетки располагаются в 3-4 ряда (общее число от 12000 до 20000). Каждая волосковая клетка имеет удлиненную форму. Один полюс клетки фиксирован на основной мембране, второй находится в полости перепончатого канала улитки. На конце этого полюса рецепторной клетки имеются волоски (30-40) на внутренней клетке они очень короткие (4-5 мкм), на наружной клетке число волосков от 65 до 120 тоньше и длиннее.
Над кортиевым органом лежит текториальная мембрана, один край которой закреплен, второй - свободен. При действии звука основная мембрана начинает колебаться, длинные волосковые клетки (стериоцилии) деформируются. Это приводит к открытию ионных каналов в мембране волосковой клетки - начинает течь калиевый ионный ток. Происходит деполяризация пресинаптического окончания волосковой клетки, что приводит к выходу в синаптическую щель нейромедиатора ( глутамата или аспартата ). Оказалось, что все стериоцилии одного рецептора связаны между собой в пучок тонкими поперечными нитями. Поэтому когда сгибаются один или несколько более тонких волосков, они тянут за собой все остальные волоски. Это вызывает генерацию электрических (импульсов) или потенциалов, а затем через синапсы - возбуждение слухового нерва, который несет информацию в ЦНС.
Механизмы восприятия звука Слуховые ощущения Слух представляет собой функцию организма, которая неразрывно связана по своему происхождению со звуком. ВЕСТИБУЛЯРНЫЙ АНАЛИЗАТОР
Вестибулярная система играет наряду со зрительной и соматосенсорной системами ведущую роль в пространственной ориентации человека. Она получает, передает и анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения, а также при изменении положения головы в пространстве. При равномерном движении или в условиях покоя рецепторы вестибулярной сенсорной системы не возбуждаются. Импульсы от вестибулорецепторов вызывают перераспределение тонуса скелетной мускулатуры, что обеспечивает сохранение равновесия тела. Эти влияния осуществляются рефлекторным путем через ряд отделов ЦНС. Проприоцептивный АНАЛИЗАТОР Двигательный (проприоцептивный) анализатор обеспечивает формирование так называемого «мышечного чувства» при изменении напряжения мышц, их оболочек, суставных сумок, связок, сухожилий. В мышечном чувстве можно выделить три составляющих: чувство положения, когда человек может определить положение своих конечностей и их частей относительно друг друга; чувство движения, когда, изменяя угол сгибания в суставе, человек осознает скорость и направление движения; чувство силы, когда человек может оценить мышечную силу, нужную для движения или удерживания суставов в определенном положении при подъеме или перемещении груза. Наряду с кожным, зрительным, вестибулярным двигательный анализатор оценивает положение тела в пространстве, позу, участвует в координации мышечной деятельности. Периферический отдел представлен проприорецепторами, расположенными в мышцах, связках, сухожилиях, суставных сумках, фасциях. К ним относятся мышечные веретена, тельца Гольджи, тельца Фатера—Пачини, свободные нервные окончания. Мышечное веретено представляет собой скопление тонких коротких поперечнополосатых мышечных волокон, которые окружены соединительнотканной капсулой. Эти волокна получили название интрафузальных в отличие от обычных мышечных волокон, которые составляют основную массу мышц и называются экстрафузальными, или рабочими, волокнами. Интрафузальные волокна расположены параллельно экстрафузальным, поэтому возбуждаются при расслаблении (удлинении) скелетной мышцы.
Рассмотрим подробно миотатический рефлекс. В мышце есть три основных типа проприорецепторов: 1) первичные окончания веретён; 2) вторичные окончания веретён; Рецепторы Гольджи.
Мышечное веретено длиной 3 мм шириной 0,3 мм состоит из интрафузальных мышечных волокон, которые окружены толстой соединительнотканной капсулой, внутри которого находится жидкость, подобная лимфе. Два толстых волокна имеют по экватору скопление ядер (ядерная сумка), и волокна называются ядерно-сумчатыми. Четыре тонких волокна имеют по экватору ядра расположенные в один ряд (ядерная цепочка), а волокна получили название ядерно-цепочечных. В разных скелетных мышцах число веретен на 1 г ткани варьирует от нескольких единиц до сотни. На интрафузальных волокнах спирально расположены чувствительные окончания афферентных волокон группы Iа — Эти волокна называются первичными афферентами. Первичные окончания (около 15мкм) -миелинизированное чувствительное нервное волокно Ia (скорость проведения 70–120 м/с) формирует первичное, или аннулоспиральное окончание, (они представляют из себя рецепторную спираль). Эти окончания возбуждаются при расслаблении или удлинении мышцы, вызванной растяжением, и реагируют на скорость удлинения экстрафузальных волокон. При механической деформации аннуло-спиральных окончаний (периферическое воздействие) рождается рецепторный потенциал, который далее преобразуется в потенциал действия и распространяется по толстым афферентным волокнам первой группы ( Iа ) к спинальным ганглиям. Далее волокна от спинального ганглия направляются к а-фазическим нейронам и от них к белым мышцам (быстрым нейромоторным единицам). Импульсация, идущая от веретен по афферентным волокнам группы Iа в спинном мозге моносинаптически возбуждает мотонейроны своей мышцы и через тормозящий интернейронтормозит мотонейроны мышцы-антагониста ( реципрокное торможение). Аннуло-спиральные терминали могут также возбуждаться (в ответ на их деформацию) при сокращении интрафузальных мышечных волокон под влиянием нервных импульсов от γ-нейронов (центральное воздействие).
К ядерно-цепочечным волокнам также подходят чувствительные окончания афферентных волокон группы II - (образовано 1–2 тонкими чувствительными нервными волокнами диаметром около 8мкм.) — так называемые вторичные окончания которые заканчиваются на них гроздьюбляшек - это колечки, розетки, утолщения терминалей. Они механически деформируются и возбуждаются под влиянием статической длины мышцы. Афферентные волокна группы II возбуждают мотонейроны мышц-сгибателей и тормозят мотонейроны мышц-разгибателей. Имеются, однако, данные, что афферентные волокна группы II, идущие от мышц-разгибателей, могут возбуждать мотонейроны своей мышцы. Потенциалы действия, пришедшие к атоническим нейронам по волокнам второй группы, далее направляются к красным мышцам (медленным нейромоторным единицам). Гамма-регуляция мышечного тонуса. Степень сокращения (напряжения) мышцы зависит от частоты импульсов, приходящих к ней от а-мотонейронов. Частота разрядов а-мотонейронов в свою очередь регулируется импульсами от проприорецепторов этой же мышцы. Веретена имеют и эфферентную иннервацию: интрафузальные мышечные волокна иннервируются аксонами, идущими к ним от γ -мотонейронов. Эти так называемые γ -эфферентные волокна подразделяют на динамические и статические. Разряды γ-мотонейронов повышают чувствительность мышечных веретён, причем динамические γ -эфференты преимущественно усиливают реакцию на скорость удлинения мышцы, а статические — на длину. Активация у-эфферентов и без растяжения мышцы сама по себе вызывает импульсацию афферентов веретен вследствие сокращения интрафузальных мышечных волокон. В результате получается, что при одинаковой длине мышцы увеличивается поток импульсов от рецепторов к а-мотонейронам, а от них - к мышце. Тем самым повышается мышечный тонус. Уровень возбуждения γ -системы тем выше, чем интенсивнее возбуждены а-мотонейроны данной мышцы, т. е. чем больше сила ее сокращения. Сами γ -мотонейроны контролируются ретикулярной формацией ствола мозга, мозжечком и корой. Благодаря сложному строению мышечные веретёна реагируют не только на периферические возбуждения (изменение длины мышцы), но и на изменения активных центральных механизмов - возбуждение γ -мотонейронов («γ -петля»). Таким образом, веретена можно рассматривать как непосредственный источник информации о длине мышцы и ее изменениях, если только мышца не возбуждена. При активном состоянии мышцы необходимо учитывать влияние у-системы. Во время активных движений γ -мотонейроны поддерживают импульсацию веретен укорачивающейся мышцы, что дает возможность рецепторам реагировать на неравномерности движения как увеличением, так и уменьшением частоты импульсации и участвовать таким образом в коррекции движений. Сухожильные рецепторы Гольджи (тельца Гольджи) находятся в сухожилиях. Это гроздьевидные чувствительные окончания, достигающие у человека 2—3 мм в длину и 1 — 1,5 мм в ширину. Тельца Гольджи, располагаясь в сухожилиях, включены относительно скелетной мышцы последовательно, поэтому они возбуждаются при ее сокращении вследствие натяжения сухожилия мышцы. Они имеют высокий порог возбуждения. Рецепторы Гольджи контролируют силу мышечного сокращения — напряжения. Идущие от этих рецепторов афферентные волокна относятся к группе Ib. На спиналь-ном уровне они через интернейроны вызывают торможение мотонейронов собственной мышцы и возбуждение мотонейронов мышцы-антагониста. Информация от мышечных рецепторов по восходящим путям спинного мозга поступает в высшие отделы ЦНС, включая кору большого мозга, и участвует в кинестезии.
Таким образом, состояние мышцы контролируется двумя типами рецепторов – мышечными веретенами, передающими информацию о длине мышцы и скорости изменения этой длины, и сухожильным рецептором, который сигнализирует о силе мышечного сокращения. В отличие от мышечных веретён рецепторы Гольджи не имеют собственной эфферентной иннервации. Их импульсы направляются в спинной мозг к вставочным тормозным нейронам. Далее происходит переключение импульсов на а-мотонейроны, которые тормозятся, в результате чего мышца расслабляется. Это называется «аутогенным торможением». Все сухожильные рефлексы начинаются с рецепторов Гольджи. Коленный рефлекс — это искусственный (сухожильный) рефлекс. Он наступает при растяжении мышечного веретена в ответ на удар молоточком. Коленный рефлекс в отличие от других рефлексов не может наступать с рецепторов Гольджи, так как для их раздражения нужна большая сила, а у мышечного веретена порог возбуждения низкий. От рецепторов возбуждение поступает в Ib, а оттуда на четырёхглавую мышцу бедра, и нога в коленном суставе выпрямляется. Тельца Фатера — Почини представляют собой инкапсулированные нервные окончания, локализуются в глубоких слоях кожи, в сухожилиях и связках, реагируют на изменения давления, которое возникает при сокращении мышц и натяжении сухожилий и связок.
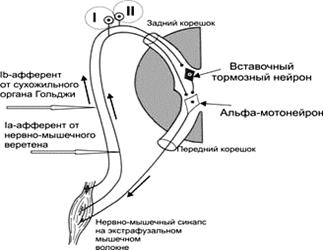
Рис. 1. Рефлексы растяжения моносинаптический (I, от мышечных веретён, приводит к сокращению той же мышцы) и полисинаптический (II) Моносинаптическая дуга. Ia–проприоцептивные нервные волокна, отходящие от мышечного веретена, входят в задний корешок спинного мозга и сразу направляются в передний рог, где и образуют синапсы с a‑мотонейронами, посылающими сигналы к мышце. Полисинаптическая дуга дополнительно включает вставочный нейрон. На рис.1. - (II) представлена дуга тормозного рефлекса, возникающего при растяжении сухожильных рецепторов Гольджи. Проводниковый отдел. Чувствительность кожи и ощущение движения обусловлены проведением в мозг сигналов от рецепторов по двум основным путям (трактам): леминисковому и спинно-таламическому, значительно различающимся по своим морфологическим и функциональным свойствам. Существует и третий путь — латеральный тракт Морина, близкий по ряду характеристик к леминисковой системе. Леминисковый путь на всех уровнях состоит из относительно толстых и быстропроводящих миелинизированных нервных волоконПервые нейроны этого пути находятся в спинномозговом узле, их аксоны в составе задних столбов восходят к тонкому (ядро Голля) и клиновидному (ядро Бурдаха) ядрам продолговатого мозга, где сигналы передаются на вторые нейроны леминискового пути. Часть волокон, в основном несущих сигналы от суставных рецепторов, оканчивается на мотонейронах сегментарного спинального уровня. Проприорецептивная чувствительность передается в спинном мозге также по дорсальному спинно-мозжечковому, спинно-цервикальному и некоторым другим путям. В продолговатом мозге в клиновидном ядре — вторые нейроны проприорецептивной чувствительности. Аксоны этих нейронов образуют медиальную петлю и после перекреста на уровне олив направляются в специфические ядра таламуса — вентробазальный ядерный комплекс. В этих ядрах концентрируются третьи нейроны леминискового пути. Их аксоны направляются в соматосенсорную зону коры большого мозга. Спинно-таламический путь. Этот путь значительно отличается от лемнискового. Его первые нейроны также расположены в спинномозговом узле, откуда они посылают в спинной мозг медленнопроводящие немиелинизированные нервные волокна. Эти нейроны имеют большие рецептивные поля, иногда включающие значительную часть кожной поверхности. Вторые нейроны данного пути локализуются в сером веществе спинного мозга, а их аксоны в составе восходящего спинно-таламического пути направляются после перекреста на спинальном уровне в вентробазальный ядерный комплекс таламуса (дифференцированные проекции), а также в вентральные неспецифические ядра таламуса, внутреннее коленчатое тело, ядра ствола мозга и гипоталамус. Локализованные в этих ядрах третьи нейроны спинно-таламического пути лишь частично дают проекции в соматосенсорную зону коры.
ЗАНЯТИЕ 31СЛУХОВАЯ СЕНСОРНАЯ СИСТЕМА
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-08-12; просмотров: 430; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 13.59.100.42 (0.029 с.) |