Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
IY. Фаза формирования (спермиогенез)Стр 1 из 7Следующая ⇒
ГАМЕТОГЕНЕЗ Развитие половых клеток называется прогенезом, или гаметогенезом, и происходит в половых железах – семенниках в мужском организме и яичнике в женском организме. Суть гаметогенеза состоит в образовании из диплоидных стволовых предшественников высокодифференцированных половых клеток – сперматозоидов и яйцеклеток, содержащих гаплоидный набор хромосом. Первичные половые клетки гоноциты (гонобласты) впервые выявляются в стенке желточного мешка (в конце третьей недели жизни эмбриона человека). Гоноциты являются индифферентыми клетками: они не разделяются на мужские и женские половые клетки. Затем гоноциты мигрируют в закладки половых желез. По пути они многократно делятся. Гоноциты дифференцируются в сперматогонии (в мужском организме) или в овогонии (в женском организме). Это процесс начинается на 6-ой недели жизни эмбриона. СПЕРМАТОГЕНЕЗ – развитиемужских половых клеток - происходит в извитых канальцах семенника (рис.1). В процессе сперматогенеза выделяют 4 фазы (периода): • размножение (пролиферация) • рост • созревание • формирование (спермиогенез). I. Фаза размножения: Сперматогенез происходит в извитых канальцах семенника. Фаза размножения, как и сперматогенез в целом, начинается с момента полового созревания. Мужские половые клетки в этой фазе представлены диплоидными клетками – сперматогониями, которые делятся путём митотических делений, обычных для всех соматических клеток. Сперматогонии располагаются в нижнем ряду сперматогенного эпителия, рядом с базальной мембраной (рис.2).
Рис.1. Схема сперматогенеза и овогенеза
Рис.2. Участок стенки извитого канальца семенника 1 – миоидные клетки стенки; 2 – базальная мембрана; 3 - сперматогонии А; 4 – сперматогонии В; 5 – сперматоцит первого порядка; 6 – сперматоцит второго порядка; 7 – сперматиды на ранней стадии развития; 8 – сперматиды в конце развития; 9 – клетка Сертоли.
Различают 3 типа сперматогоний (рис.2): • тёмные А сперматогонии – истинные стволовые клетки, устойчивы к действию вредных факторов, делятся редко; • светлые А сперматогонии – полустволовые клетки, способны к частым митотическим делениям (результат деления: образуются две А- сперматогонии, либо одна А и одна В-сперматогонии).
• светлые В сперматогонии, коммитированные клетки, которые также проходят несколько митотических циклов и далее вступают в фазу роста. II. Фаза роста В сперматогонии вступают в профазу первого мейотического деления, перемещаясь из базального слоя по направлению к просвету канальца. На этой стадии развития половые клетки называются сперматоцитами первого порядка (первичными сперматоцитами). Рост характеризуется увеличением объёма сперматоцитов приблизительно в 4 раза. Профаза первого деления мейоза довольно длинная: у человека она продолжается около 22 суток. III. Фаза созревания В результате первого деления мейоза (редукционного) образуются сперматоциты второго порядка (вторичные сперматоциты), которые быстро вступают во второе деление мейоза (эквационное). В результате второго мейотического деления образуются клетки, которые называются сперматиды. Сперматиды имеют гаплоидный набор хромосом. Начиная со сперматогоний типа В, дочерние клетки при делении не отделяются полностью друг от друга, а остаются связанными с помощью цитоплазматических мостиков, образуя синцитий. Лишь зрелые сперматозоиды отделяются от синцития. IY. Фаза формирования (спермиогенез) Фаза формирования - самая продолжительная фаза сперматогенеза (около 50 суток). В процессе формирования зрелых сперматозоидов из сперматид происходят следующие изменения в строении клетки (рис.3):
Рис.3. Важнейшие изменения, происходящие в сперматидах в период формирования
• конденсация ядерного хроматина; конденсация хроматина приводит к образованию суперкомпактного ядра, что увеличивает мобильность сперматозоида и защищает геном от повреждения; в укладке хроматина ядра спермия принимают особые гистоны и белки протамины; • образование акросомы из элементов комплекса Гольджи; акросома - специальный тип лизосомы, содержит гидролитические ферменты (такие как гиалуронидаза, нейраминидаза, кислая фосфатаза, протеазы), необходимые для разрушения оболочек яйцеклетки; акросома в виде «шапочки» покрывает переднюю половину ядра; наружная мембрана акросомы соприкасается с клеточной мембраной, а внутренняя – с мембраной ядра;
• центриоли перемещаются к противоположному полюсу ядра; от дистальной центриоли образуется осевая нить (аксонема) жгутика. • митохондрии в виде спирали окружают часть аксонемы, образуя утолщенный участок – промежуточный отдел; митохондрии обеспечивают энергию для движения сперматозоида; • цитоплазма редуцируется до минимума; • после сброса излишка цитоплазмы (которое называют остаточным тельцем), сперматозоиды отделяются от общей ассоциации клеток - синцития - и становятся свободными. В результате мейоза в сперматогенезе из одной исходной сперматогонии образуются 4 сперматозоида. Рис.4. Схема строения сперматозоида ОВОГЕНЕЗ Образование женских половых клеток – овогенез – происходит в яичнике. Овогенез включает 3 фазы (рис.1): • размножение • рост • созревания I. Фаза размножения В отличие от сперматогенеза, фаза размножения в овогенезе протекает исключительно в эмбриональный период развития. Овогонии делятся путём митозов. Большинство образующихся дочерних клеток погибает (этот процесс называется атрезия), часть овогоний вступает в фазу роста (в профазу первого деления мейоза). Митотические деления овогоний прекращаются к седьмому месяцу эмбриогенеза. При рождении все яйцеклетки в яичнике являются овоцитами первого порядка (первичными овоцитами), которые блокированы в профазе первого деления мейоза. II. Фаза роста Фаза роста соответствует жизни первичного овоцита, находящегося в профазе первого деления мейоза. Фаза роста делится на два периода: малого роста (до полового созревания) и большого роста. К моменту рождения в яичнике насчитывается около 2 млн первичных овоцитов; к периоду полового созревания большинство из них погибает, остаётся около 400 тысяч яйцеклеток. Рост овоцитов происходит в фолликулах – структурах, в которых овоцит окружен эпителиальными фолликулярными клетками. В период малого роста имеется два типа фолликулов:
Вступление овоцитов в период большого роста происходит только под воздействием гормонов (фолликулостимулирующего гормона); этому периоду оогенеза соответствуют растущие (вторичные и третичные) фолликулы. Период роста в овогенезе очень длительный, может продолжаться от 12 до 50 лет. III. Фаза созревания Первое деление мейоза завершается только перед овуляцией (овуляция -разрыв зрелого третичного фолликула и выброс яйцеклетки из яичника в брюшную полость). В результате первого деления мейоза образуется овоцит второго порядка (вторичный овоцит). Овоцит второго порядка вступает во второе деление мейоза, но это деление блокируется на стадии метафазы и завершается только при условии оплодотворения. За один менструально-овариальный цикл (~ 28 дней) происходит овуляция обычно одной яйцеклетки. Таким образом, за репродуктивный период жизни женщины (~ 30-40 лет) только около 450 яйцеклеток достигают зрелости – готовности к оплодотворению.
Мейотическое деление в овогенезе отличается тем, что протекает с неравной цитотомией: в мейозе I хромосомы поровну распределяются между дочерними клетками, но только одна из клеток (вторичный овоцит) сохраняет почти всю цитоплазму; вторая клетка (первое полярное, или редукционное, направительное тельце) имеет очень мелкие размеры, небольшое ядро, минимум цитоплазмы. Редукционное тельце, которое образуется в результате первого деления мейоза, при втором делении также делится, давая два дочерних тельца. Таким образом, в результате мейоза в овогенезе образуется лишь одна зрелая яйцеклетка и три редукционных тельца. Зрелая яйцеклетка млекопитающих – крупная (диаметр 100-200 мкм) в сравнении со сперматозоидом, неподвижная клетка. Особенность строения яйцеклетки – отсутствие центриолей и наличие характерных трофических включений – желточных (вителлиновых) гранул. В цитоплазме яйцеклетки хорошо развита система белкового синтеза. По периферии яйцеклетки, под клеточной мембраной, равномерно распределены кортикальные гранулы, содержащие ферменты (гидролазы), необходимые для формирования оболочки оплодотворения после проникновения сперматозоида в яйцеклетку. При овуляции яйцеклетка окружена (рис.5): • прозрачной оболочкой (zona pellucida), содержащей густую сеть тонких нитей, состоящих из гликопротеинов (фракции ZP1, ZP2, ZP3); • фолликулярными клетками, образующими лучистый венец (corona radiata).
Млекопитающего 1 – ядро; 2 – микроворсинки яйцеклетки; 3 – фолликулярные клетки лучистого венца; 4 – отростки фолликулярных клеток; 5 – кортикальные гранулы; 6 – желточные включения; 7 – прозрачная оболочка
Основные различия в процессах сперматогенеза и овогенеза представлены в таблице 1. Таблица 1 ОПЛОДОТВОРЕНИЕ Оплодотворение -цепь событий, завершающаяся процессом слияния сперматозоида и яйцеклетки, что приводит к образованию одноклеточного зародыша – зиготы. При овуляция вторичный овоцит, окруженный прозрачной оболочкой и лучистым венцом, выбрасывается из яичника в брюшную полость и попадает в ампулу маточной трубы, в которой может произойти оплодотворение. Если этого не происходит в течение первых 24 часов после овуляции, овоцит дегенерирует. В эякуляте человека содержится 3х108 сперматозоидов. В женских половых путях они сохраняют способность к оплодотворению до двух суток. Только около 200 из них достигают ампулярной части маточной трубы (яйцевода), где и происходит оплодотворение.
Условно в процессе оплодотворения выделяют следующие ФАЗЫ: • дистантное взаимодействие; • контактные взаимодействия; • фаза синкариона, слияние пронуклеусов и образование зиготы. Процесс оплодотворения длится от 12 до 24 часов. ДИСТАНТНОЕ ВЗАИМОДЕЙСТВИЕ Сперматозоиды направленно мигрируют к воронке маточной трубы, где и происходит встреча с яйцеклеткой. При этом важную роль играют сокращения мышечных волокон стенки маточной трубы и хемотаксис – направленная миграция по градиенту химических веществ - хемоаттрактантов. Хемоаттрактанты попадают в маточную трубу в составе фолликулярной жидкости лопнувшего фолликула, секретируются яйцеклеткой (гиногамоны) и фолликулярными клетками лучистого венца. В женских половых путях под влиянием факторов женского организма (слизистая секреция, рН и др.) происходит капацитация – физиологический процесс приобретения сперматозоидом способности к оплодотворению: окончательное созревание и активация. Во время капацитации происходят: • удаление гликопротеинов с цитолеммы головки сперматозоида; • перестройка цитолеммы (она становится более «текучей», «открытой» для проникновения ионов Са), что приводит к гиперактивации и усилению подвижности сперматозоида. Для завершения капацитации требуется около 7 часов. Капацитацию можно воспроизвести in vitro. КОНТАКТНЫЕ ВЗАИМОДЕЙСТВИЯ Для того чтобы произошло оплодотворение, сперматозоид должен последовательно преодолеть три барьера: • лучистый венец, из нескольких слоёв рыхло расположенных фолликулярных клеток; • прозрачную оболочку; • плазматическую мембрану (оолемму) яйцеклетки. Сперматозоид довольно легко преодолевает лучистый венец, поскольку между его клетками нет прочных связей. Более существенным барьером для сперматозоида является прозрачная оболочка. При контакте с прозрачной оболочкой сначала происходит связывание и прикрепление сперматозоида с рецепторами прозрачной оболочки ZP2, ZP3. Благодаря рецепторам это прикрепление является видоспецифическим: сперматозоид и яйцеклетка должны обязательно принадлежать к одному виду млекопитающего. После прочного связывания сперматозоида с ZP3 наступает акросомная реакция: мембрана акросомы во многих местах сливается с плазматической мембраной спермия в передней части головки, ферменты акросомы выделяются и расщепляют компоненты прозрачной оболочки, обеспечивая проникновение сперматозоида через образовавшийся канал к плазматической мембране яйцеклетки. Активные движения хвостика позволяют сперматозоиду мигрировать через прозрачную оболочку за 5-10 мин. Касание одного из сперматозоидов оолеммы (плазматической мембраны яйцеклетки) приводит к образованию на поверхности воспринимающего бугорка. Оолемма содержит систему рецепторов, благодаря которым становится возможным слияние плазматических мембран головки сперматозоида и яйцеклетки, в результате чего головка спермия внедряется в ооплазму (в ооплазму входят только ядро сперматозоида и его центриоли).
Одновременно с началом взаимодействия двух гамет в яйцеклетке происходит кортикальная реакция, запускаемая быстрым повышением концентрации ионов кальция. Содержимое кортикальных гранул яйцеклетки выделяется в перивителлиновое пространство (пространство между оолеммой и прозрачной оболочкой). Ферменты кортикальных гранул расщепляют рецепторы ZP2 и модифицируют ZP3, при этом блестящая оболочка теряет способность связывать другие спермии и блокирует акросомную реакцию других спермиев – формируется твердая оболочка оплодотворения, непреодолимая для спермиев. Оолемма также приобретает новые свойства, в том числе отрицательный заряд, отталкивающий спермии, в результате чего другие сперматозоиды уже не могут проникнуть в ооплазму. Таким образом, оболочка оплодотворения обеспечивает блок полиспермии. Предупреждение полиспермии создаёт условия для наступления нормальной беременности. ФАЗА СИНКАРИОНА Проникновение сперматозоида в яйцеклетку запускает завершение второго мейотическогоделения овоцита. В телофазе II формируются ядерные оболочки и образуются два гаплоидных ядра. Одно из них, окруженное тонким слоем цитоплазмы отделяется от овоцита в виде второго полярного тельца. Другое ядро, оставшееся в овоците, теперь называется женским пронуклеусом. Ядро проникшего сперматозоида также претерпевает ряд преобразований и набухает, формируя мужской пронуклеус. Оба пронуклеуса и центриоли сперматозоида перемещаются к центру ядра. В каждом из них начинается подготовка к митозу (конденсация и перестройка хроматина), происходит удвоение ДНК. После сближения пронуклеусов их ядерные оболочки исчезают, а уже удвоенные хромосомы оказываются лежащими в цитоплазме. Формируется общее митотическое веретено и начинается первое деление дробления. Результат оплодотворения – образование зиготы. ЗИГОТА -оплодотворенная яйцеклетка и (или) новый организм, возникающий при слиянии двух гамет на одноклеточной стадии развития.
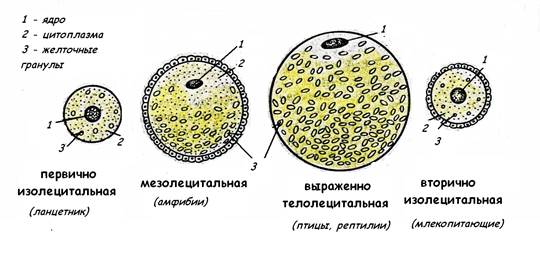
Рис.6. Последовательность контактных взаимодействий в процессе оплодотворения ТИПЫ ЯЙЦЕКЛЕТОК Размеры и строение яйцеклеток различных представителей животного мира обусловлены количеством накопленных в цитоплазме питательных веществ, необходимых для формирования и роста зародыша. Эти питательные вещества содержатся в виде желточных включений (комплекса белков, липидов и углеводов). Количество желтка в цитоплазме яйцеклетки различно в зависимости от условий развития. Поскольку желток тормозит или даже препятствует делению цитоплазмы, количество и распределение желтка определяет тип делений дробления и образования бластулы. Таким образом, классификации яйцеклеток основывается на количестве и характере распределения желточных включений в цитоплазме яйцеклетки. У различных животных в зависимости от количества питательных веществ в цитоплазме яйцеклетки могут быть (рис.7): • алецитальными – практически не содержат желтка; • олиголецитальными –с малым количеством желтка; • полилецитальными – с большим количеством желтка. В олиголецитальных яйцеклетках небольшое количество желтка равномерно распределено в цитоплазме; такие яйцеклетки называются изолецитальными. Полилецитальные яйцеклетки содержат асимметричное скопление желтка у одного из полюсов яйцеклетки, называемого вегетативным, ядро располагается ближе к противоположному анимальному полюсу. Эта полярность в распределении желтка может быть более умеренной - в мезолецитальных яйцеклетках (у амфибий) или резко выраженной – в телолецитальных яйцеклетках (у рептилий, птиц). В центролецитальных яйцеклетках желток равномерно распределён по всей цитоплазме, за исключением её периферического слоя (у многих членистоногих).
Рис.7. Типы яйцеклеток
У примитивных хордовых типа ланцетника, которые ведут водный способ существования и имеют стадию личинки – яйцеклетка первично изолецитального типа. У некоторых рыб и амфибий, с развитием через личиночную стадию, яйцеклетки имеют умеренный запас питательных веществ – это мезолецитальные яйцеклетки. У рептилий и птиц, развитие которых происходит на суше и для развития требуется большой запас питательных веществ, яйцеклетки крупные, богатые желтком,- резко телолецитальные яйцеклетки. В филогенезе предки высших млекопитающих также имели телолецитальные яйцеклетки, но в связи с переходом на внутриутробное развитие плода, к которому все необходимые вещества поступают из крови матери через плаценту, количество желтка было редуцировано, и яйцеклетка высших млекопитающих, включая яйцеклетку человека, относится к вторично изолецитальному типу. ДРОБЛЕНИЕ Дробление – стадия развития зародыша, которая следует за оплодотворением и включает в себя ряд последовательных митотических делений без последующего роста дочерних клеток, называемых делениями дробления. За время короткой интерфазы происходит в основном только удвоение ДНК. РНК и белки практически не синтезируются (G1 фаза клеточного цикла отсутствует). Клетки, образующиеся при делениях дробления, называются бластомерами. Дробление происходит до тех пор, пока бластомеры не достигнут размеров, обычных для соматических клеток данного организма. В результате дробления возникает многоклеточный зародыш – бластула. У разных видов животных процессы дробления характеризуются рядом видовых особенностей, обусловленных строением яйцеклетки: характер дробления определяется количеством желтка и разным распределением его в цитоплазме яйцеклетки. Дробление может быть: • полным (голобластическим), когда весь объём ооцита делится на бластомеры; • неполным (меробластическом, частичным), когда делится только часть ооцита, его анимальный полюс, относительно свободный от желтка (характерно для резко телолецитальной яйцеклетки) Кроме того, дробление может быть: • равномерным – если все бластомеры, образующиеся при дроблении одинаковы; • неравномерным – если бластомеры, образующиеся при дроблении имеют неравные размеры. Рис.8. Ранние стадии развития ланцетника
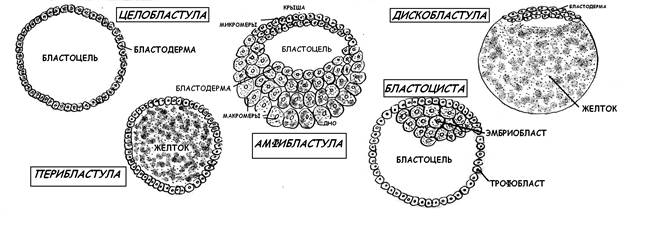
В результате полного неравномерного дробления мезолецитальной яйцеклетки амфибий (лягушки) образуется амфибластула (рис.9) : • на анимальном полюсе, которое содержало небольшое количество желтка, клеточные деления происходят с большой скоростью и образуются мелкие бластомеры – микромеры; • на вегетативном полюсе, богатом желтком, клетки делятся медленно и образуются крупные бластомеры – макромеры; • бластодерма (стенка бластулы) многослойная; • бластоцель смещён к анимальному полюсу; • анимальный полюс с многочисленными микромерами называется крышей бластулы, вегетативный с с макромерами образует дно; между ними краевая зона.
Рис.9. Ранние стадии эмбриогенеза амфибий Тип дробления у птиц с полилецитальной резко телолецитальной яйцеклеткой - неполное (частичное) дискоидальное: деление ограничено небольшим участком на анимальном полюсе, остальная часть желтка остаётся нераздробленной. В результате такого типа дробления образуется дискобластула (рис.10): • дискобластула имеет форму круглой пластинки (диска) на анимальном полюсе; бластодерма многослойная; • щелевидный бластоцель находится между бластодермой и синцитиальном слоем на поверхности желтка.
Рис.10. Типы бластул Характер дробления вторично изолецитальной яйцеклетки человека – полное неравномерное асинхронное. Образующая бластула на ранней стадии носит название морула (до стадии 64 бластомеров), а после появления полости – бластоцеля – бластоциста (рис.10). ГАСТРУЛЯЦИЯ Гаструляция – это следующий за дроблением период развития зародыша. Скорость клеточных делений снижается, интерфаза между митозами удлиняется, а сами клетки начинают активно синтезировать РНК и белки. Главная особенность этого периода – направленные активные перемещения делящихся клеток (клеточных потоков), которые приводят к глубокой перестройке зародыша и в конечном итоге к образованию трёх зародышевых листков: • эктодерма – наружный зародышевый листок; • энтодерма – внутренний зародышевый листок; • мезодерма - средний зародышевый листок, лежащий между экто- и энтодермой Способы гаструляции у животных разных систематических групп сильно варьируют. Описано четыре основных типа морфогенетических движений гаструляции (рис.11): • инвагинация; • эпиболия; • деламинация; • иммиграция
Рис.11. Основные механизмы гаструляции
Инвагинация – это впячивание бластодермы вегетативного полюса бластулы в полость бластоцеля, при этом образуется первичная кишка с полостью – гастроцелем, который сообщается с наружной средой отверстием – бластопором (первичным ртом). Инвагинация происходит при гаструляции у примитивных хордовых, таких как Ланцетник (рис.8). Эпиболия – перемещение активно делящихся мелких клеток (микромеров) анимальной части бластодермы в направлении вегетативного полюса. Микромеры обрастают крупные клетки (макромеры) вегетативного полюса, и макромеры погружаются внутрь зародыша. Эпиболия, как тип движений гаструляции, характерна для эмбриогенеза амфибий. Деламинация – расщепление (расслоение) бластодермы на два пласта: наружный – эпибласт и внутренний – гипобласт. Иммиграция – перемещение отдельных клеток из бластодермы внутрь зародыша. Если небольшие участки определенной области поверхности бластулы окрасить прижизненным красителем, можно проследить дальнейшую судьбу, дифференцировку клеток этого участка. Экспериментально показано, что клетки анимального полюса – клетки будущей эктодермы. По результатам таких исследований можно составить карту презумптивных зон. ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ ПТИЦ Тип яйцеклетки у птиц – полилецитальная, резко телолецитальная яйцеклетка. Тип дробления зиготы птиц- неполное (частичное, меробластическое) дискоидальное. Дробление ограничено лишь малой частью зиготы на анимальном полюсе (дробится лишь 1/500 часть зиготы), остальная часть зиготы не подвергается делению и представляет собой нераздробившийся желток. При дроблении возникают бластомеры различной величины, и в конечном итоге формируется дискобластула: бластомеры располагаются в форме диска, лежащего на массе нераздробившегося желтка (рис.10). Центральная часть дискобластулы называется зародышевым диском (бластодиском). Гаструляция в дискобластуле происходит в две фазы:
Первая фаза гаструляции- деламинация - заключается в расщеплении клеточной массы бластодиска на 2 листка: гипобласт (первичная энтодерма) и эпибласт (первичная эктодерма ) (Рис.12).
Рис.12. Деламинация в эмбриогенезе птиц
4. нераздробившийся желток.
Клетки внутреннего листка - гипобласта - смыкаются в правильную однослойную пластинку. Наружный слой бластодиска – эпибласт – представляет собой клеточный пласт из более высоких призматических клеток. Центральный утолщенный участок бластодиска, состоящий из эпибласта и гипобласта, называется зародышевым щитком. Из зародышевого щитка образуется тело самого зародыша. Вторая фаза гаструляции – иммиграция связана со сложным перемещением клеток эпибласта сначала к заднему (хвостовому) концу зародыша (рис.13,А). Клетки эпибласта движутся двумя потоками по краям зародышевого диска, оба потока сталкиваются в хвостовом отделе у медианной (средней) линии, сливаются и затем направляются вперёд, к головному концу, образуя утолщенной клеточный тяж по средней линии – первичную полоску. Второй поток клеток (более медленный) распространяется со стороны головного конца зародыша и направляется по средней части эпибласта навстречу первому потоку. Место встречи двух потоков - первичный (гензеновский) узелок. Вдоль первичной полоски образуется углубление – первичная бороздка. Из первичной полоски клетки выселяются и вначале встраиваются в первичную энтодерму, раздвигая клетки гипобласта. Так формируется энтодерма зародыша. Далее клетки мигрируют из первичной полоски в обе стороны от неё, и встраиваются между первичной эктодермой и энтодермой, образуя средний зародышевый листок - мезодерму. В процессе гаструляции происходит также образование мезенхимы – эмбрионального зачатка соединительной ткани. Мезенхима выселяется из всех зародышевых листком, главным образом из мезодермы.
Рис.13. Иммиграция в эмбриогенезе птиц 1. первичный (гензеновский) узелок; 2. первичная ямка; 3. первичная полоска; 4. первичная бороздка; 5. эпибласт; 6. гипобласт; 7. клетки, выселяющиеся из первичной полоски; 8. мезодерма; 9. зародышевая энтодерма, замещающая гипобласт.
Из материала первичного узелка через первичную ямку мигрируют клетки, образующие клеточный тяж по средней линии зародыша – хордальный отросток (нотохорд). Из зародышевых листков и мезенхимы происходит формирование зачатков всех тканей (гистогенез) и органов (органогенез) развивающегося организма. Вначале закладываются осевые зачатки органов, которые располагаются по оси симметрии тела. Комплекс осевых органов включает: • нервную трубку • хорду • параксиальную мезодерму • кишечную трубку. Нервная трубка формируется из первичной эктодермы под влиянием сигнальных молекул, вырабатываемых клетками хорды, таким образом хорда является первичным индуктором нейроэктодермы. Процесс образования нервной трубки называется нейруляцией ( рис.14 ). В зоне образования нервной трубки в эктодерме вначале появляется утолщение – нервная пластинка. В нервной пластинке образуется углубление – нервный желобок, а по краям желобка утолщения – нервные складки. При смыкании нервных складок формируются нервная трубка и нервные валики (нервные гребни).
Рис.14. Схема нейруляции Из нервной трубки образуется центральная нервная система(головнойи спинной мозг). Клетки нервного гребня активно мигрируют и дают начало структурам периферической нервной системы и некоторым другим структурам (мозговое вещество надпочечников, эндокриноциты диффузной эндокринной системы, клетки-меланоциты и др. – табл.5). Та часть эпибласта, которая остаётся после выселения клеток из первичной полоски и нейруляции, образует зародышевую эктодерму (эпидермальную эктодерму ). Зародышевая эктодермаявляется источником развития эпителия кожного покрова и его производных. В мезодерме дифференцируются следующие части (рис.15): · сомиты (параксиальная мезодерма), которые располагаются справа и слева от нервной трубки и хорды; · нефротомы (сегментные ножки, промежуточная мезодерма), лежащие латеральнее сомитов; · спланхнотомы ( латеральная мезодерма), наиболее латерально расположенные участки мезодермы.
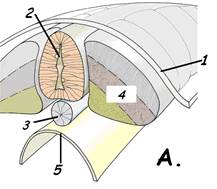
Рис.15. Зародыш курицы на этапе образования комплекса зачатков осевых органов. В.Сагиттальный срез 1.кожная эктодерма; 2.нервная трубка; 3.хорда; 4.мезодерма (сомит); 4.1. сомит (параксиальная мезодерма); 4.2.нефротом (промежуточная мезодерма, сегментные ножки); 4.3. спланхнотом (латеральная мезодерма); 4.3.1. – париетальный листок спланхнотома; 4.3.2. висцеральный листок мезодермы; 5.энтодерма; 6.целом;; 7.кровеносный сосуд.
Сомиты и нефротомы представляют собой сегментированную часть мезодермы (рис 15А). В дальнейшем сомиты далее дифференцируются на:
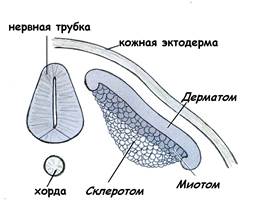
· дерматом, из которого формируется соединительнотканная часть кожи – дерма; · миотом, из которого развивается скелетная мышечная ткань; · склеротом – источник развития тканей осевого скелета (рис.16).
Рис.16. Дифференцировка сомита Нефротом служит источником для развития мочевыделительной и половой систем. Спланхнотомы не сегментируются и расслаиваются на два листка – париетальный листок, прилежащий к эктодерме (соматическаямезодерма), и висцеральный листок, прилежащий к энтодерме (спланхническаямезодерма). Между париетальным и висцеральным листками формируется целомическая полость. Из листков спланхномезодермы происходит образование эпителия серозных оболочек, выстилающих брюшину, плевру, перикард.
Из мезенхимы развиваются: · кровь и лимфа; · сосуды; · кроветворные органы; · соединительная ткань; · гладкая мышечная ткань. Из энтодермы образуется эпителий среднего отдела кишечной трубки, крупных желез, связанных с пищеварительной трубкой (печень, поджелудочная железа), эпителий лёгкого, некоторых эндокринных желез (табл.4). ВНЕЗАРОДЫШЕВЫЕ (ПРОВИЗОРНЫЕ) ОРГАНЫ обеспечивают эмбриону условия для нормального развития и жизнедеятельности. Они функционируют в течение всего эмбриогенеза и исчезают по окончании эмбрионального развития, когда зародыш приобретает способность к самостоятельному существованию. Внезародышевые органы формируются из внезародышевых частей зародышевых листков. Внезародышевые органы у птиц: · амнион; · серозная оболочка; · желточный мешок; · аллантоис. Зародыш птиц первое время распластан из поверхности желтка, но в дальнейшем формируется туловищная складка. При этом все четыре листка (эктодерма, энтодерма, париетальный и висцеральный листки мезодермы) подгибаются под зародыш, отделяя его от внезародышевых органов (рис.17). Амниотические складки, образованные из внезародышевых частей эктодермы и париетального листка мезодермы, приподнимаются сначала над головным концом, а затем над всем телом зародыша (рис.17). Эктодерма одной амниотической складки срастается с эктодермой другой складки. Париетальные листки мезодермы обеих складок также срастаются в единый слой. При этом формируются заполненный жидкостью пузырь – амнион, стенка которого образована амниотической оболочкой, состоящей из эктодермы, выстилающей его изнутри, и париетального листка мезодермы, лежащего снаружи. Эктодермальный амниотический эпителий секретирует амниотическую жидкость, обеспечивая водную среду для развития зародыша и защищая его от механических повреждений. Желточный мешок образуется из внезародышевых частей энтодермы и висцерального листка мезодермы, которые постепенно обрастают поверхность желтка (рис.18). Желточный мешок отделяется от первичной кишки в процессе формирования туловищной складки. Желток переводится в растворимую форму под действием ферментов, которые продуцируются энтодермальными клетками (клетками желточной энтодермы), и используется зародышем для питания. По мере развития зародыша и потребления питательных веществ объём желточного мешка редуцируется. Серозная оболочка образована внезародышевыми частями париетального листка мезодермы (изнутри) и эктодермы (обращена наружу). Серозная оболочка обрастает весь зародыш и выполняет функцию дыхания (газообмена). Аллантоис возникает в виде полого выроста вентральной стенки задней части кишечной трубки и состоит из энтодермы зачатка задней кишки и покрывающего его снаружи висцерального листка мезодермы с сосудами. Аллантоис постепенно разрастается, занимая всё больший объём, приближается к серозной оболочке и снабжает её сосудами.
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-08-10; просмотров: 1205; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.19.30.232 (0.139 с.) |
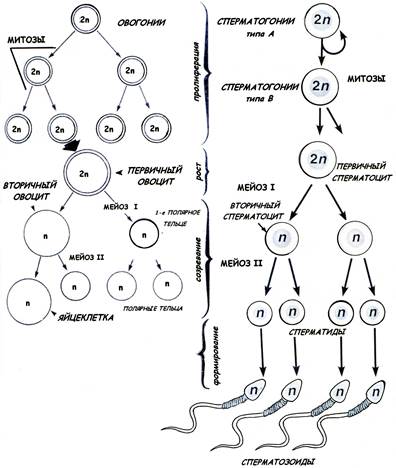
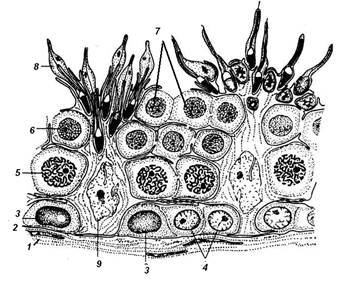

 Рис.5. Строение яйцеклетки
Рис.5. Строение яйцеклетки