
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Типы мышечной ткани: морфо-функциональные различияСтр 1 из 6Следующая ⇒
Литература основная
Типы мышечной ткани: морфо-функциональные различия
На основании чего мышечные ткани разделили на поперечнополосатую и гладкую? Важным морфологическим признаком расслабленного поперечнополосатого мышечного волокна является наличие в цитоплазме поперечной исчерченности (рис. 709250002).
Рис. 709250002. Микрофотография волокон скелетной мышцы. Хорошо видна поперечная исчерченность мышечных волокон.
Причём, поперечная исчерченность (чередование тёмных и светлых тонких полосок) - четкая. Продольная исчерченность – нечёткая. Поперечная исчерченность обусловлена упорядоченным расположением элементов актомиозинового хемомеханического преобразователя. Продольная – параллельной ориентацией миофибрилл вдоль оси мышечного волокна.
Морфо‑функциональные различия типов мышечной ткани Миокард по своим морфологическим и физиологическим характеристиками занимает промежуточное положение между скелетной и гладкой мышцей, поэтому принято сравнивать 3 типа мышц (рис. 709250936): 1. Поперечнополосатую скелетную мышцу 2. Поперечнополосатую сердечную мышцу 3. Гладкую мышцу
Рис. 709250936. Основные типы мышечных волокон. 1 - скелетный миоцит (волокно поперечнополосатой скелетной мышцы), 2 – миокардиоцит, 3 - гладкий миоцит.
Саркомер
Тёмная полоска носит название А-диска, светлая - I‑диска. А-диск в центре более светлый и эта область называется Н-полоской. Края А-диска более тёмные называются S-зоной.
Выяснилось, что I-диск в основном состоит из актиновых нитей, H‑полоска – из миозиновых, S‑зона из актиновых и миозиновых. На основании этих данных предложена схема взаимного расположения актиновых и миозиновых нитей (рис. 210211130).
Актиновые нити крепятся на Z‑мембране, миозиновые – на М‑мембране. Участок между миофибриллы между Z‑мембранами называется саркомером (рис. 210211130). Саркомер принято считать структурно-функциональной единицей (СФЕ) миофибриллы скелетных и сердечных миоцитов.
. Рис. 709261321. Серийные срезы скелетной мышцы. А – активность АТФазы: тёмные волокна – с быстрым миозином (1, 2); светлые волокна – с медленным миозином (3). B – активность СДГ: тёмные волокна – окислительные (2, 3); светлые волокна - гликолитические (1).
Оксидативные[Мф19] и гликолитические мышечные волокна
Единственным непосредственным источником энергии для мышечного сокращения является АТФ. Мышечные волокна используют два основных пути образования АТФ - окислительный и гликолитический. [i] Оксидативные [j] мышечные волокна имеют высокой уровень активности окислительных ферментов (например, сукцинатдегидрогеназы[Мф20] - СДГ) (рис. 709261321 B) и относительно низкий уровень активности гликолитических ферментов.
Рис. 709261522. Поперечные срезы скелетной мышцы. Окрашены капилляры, окружающие мышечные волокна (в основном оксидативного типа).
Рис. 709251102. Поперечные срезы скелетной мышцы. Окрашены митохондрии, которые расположены в большом количестве в волокнах оксидативного типа.
Гликолитические мышечные волокна имеют высокой уровень активности гликолитических ферментов (например, фосфофруктокиназы[Мф21] - ФФК) и относительно низкий уровень активности окислительных ферментов (рис. 709261321 B). Оксидативные мышечные волокна небольшого диаметра, окружены большим количеством капилляров (рис. 709261522), содержат много митохондрий (рис. 709251102). Гликолитические мышечные волокна большего диаметра, окружены небольшим количеством капилляров (рис. 709261522), содержат немного митохондрий (рис. 709251102).
Оксидативные мышечные волокна содержат много миоглобина и мало гликогена, поэтому имеют красный цвет и имеют второе название – красные. Гликолитические мышечные волокна содержат много гликогена и мало миоглобина, поэтому они бледны и имеют второе название – белые.
Двигательная единица
Мышечные волокна не являются функциональной единицей скелетной мускулатуры. Эту роль выполняет нейромоторная, или двигательная единица которая включает мотонейрон и группу мышечных волокон, иннервируемых разветвлениями аксона этого мотонейрона, расположенного в ЦНС Число мышечных волокон, входящих в состав двигательной единицы, различно (табл. 709261659) и зависит от функции, которую выполняет мышца в целом.
Таблица 709261659. Число мышечных волокон в двигательной единице различных мышц
В мышцах, обеспечивающих наиболее точные и быстрые движения, двигательная единица состоит из нескольких мышечных волокон, в то время как в мышцах, участвующих в поддержании позы, двигательные единицы включают несколько сотен и даже тысяч мышечных волокон.
Все мышечные волокна каждой двигательные единицы (ДЕ) относятся к одному типу. Поэтому ДЕ можно относить к типам I, IIa, IIb (рис. 709261947).
В мышце волокна разных ДЕ расположены вперемежку.
Композиция скелетных мышц Каждая мышца уникальна по спектру входящих в её состав типов мышечных волокон. В зависимости от преобладания в мышцах конкретного типа мышечных волокон скелетные мышцы относят к «красным» и «белым» либо «быстрым» и «медленным». Соответственно, общая физиологическая характеристика мышцы – сила, скорость сокращения, выносливость – в большой мере определяется процентным соотношением в мышце типов волокон. Этот спектр генетически детерминирован (отсюда практика типирования мышечных волокон при отборе спортсменов-бегунов — спринтеров и стайеров).
8. Механизм мышечного сокращения и расслабления[Мф23]
Модель скользящих нитей
При микроскопии миофибрилл в расслабленном состоянии и состоянии сокращения было отмечено, что при сокращении длина А-диска не меняется, а I-диск и H‑полоска уменьшаются и даже исчезают (рис. 709240129).
Отсюда был сделан вывод, что тонкие и толстые нити при сокращении не изменяют значимо свою длину, а скользят относительно друг друга (рис. 709240133, 709240134).
Рис. 709270802. Рабочий цикл миозиновых мостиков поперечнополосатого миоцита.
Головка миозина совершает около пяти циклов в секунду. Когда одни головки миозина толстой нити производят тянущее усилие, другие в это время свободны и готовы вступить в очередной цикл. Следующие друг за другом гребковые движения стягивают тонкие нити к центру саркомера. Скользящие тонкие нити тянут за собой Z-линии, вызывая сокращение саркомера. Поскольку в процесс сокращения практически одномоментно вовлечены все саркомеры мышечного волокна, происходит его укорочение[Мф36]. До тех пор, пока в межфибриллярном пространстве имеется Ca2+, циклы будут повторяться. Это быстрое прикрепление, разрушение и новое прикрепление поперечных мостиков происходит повторно с большой скоростью и во многих участках миофиламентов во время мышечного сокращения и прекращается, когда Ca2+ уходит из межфибриллярного пространства в цистерны или когда молекулы АТФ образуются не столь быстро, чтобы полностью обеспечивать «зарядку» энергией поперечные мостики. Напряжение, развиваемое мышечным волокном, зависит от числа замкнутых поперечных мостиков, а скорость развития напряжения — от числа замкнутых поперечных мостиков, образуемых в единицу времени, т.е. от скорости их прикрепления к актину. По-видимому, даже при максимальном возбуждении только часть поперечных мостиков эффективно связана с актиновыми миофиламентами. При повышении скорости укорочения мышцы число прикрепленных поперечных мостиков в каждый момент времени уменьшается. Это и объясняет падение силы сокращения мышцы с увеличением скорости ее укорочения[V.G.37].
Если же мышца не укорачивается (при изометрическом сокращении), напряжение в мышечных волокнах создается за счет повторного прикрепления поперечных мостиков на одних и тех же фиксированных участках актиновых нитей. В этом случае никакой внешней работы не совершается, а энергия (АТФ) расходуется на поддержание напряжения мышцы. Тяга, развиваемая сократительными элементами в миофибриллах, передается сарколемме и через нее — на сухожилия мышц. Период химико-механических изменений в сократительном миофибриллярном аппарате мышечного волокна, на протяжении которого в нем возникает и поддерживается активная механическая тяга, определяющая процесс сокращения, называется периодом активного состояния.
Расслабление Режимы мышечного сокращения
Приложение 709261417. Энергетика мышечного сокращения
Единственным прямым (непосредственным) источником энергии для мышечного сокращения служит аденозинтрифосфат (АТФ). При гидролизе АТФ образуется аденозиндифосфат (АДФ), отщепляется фосфатная группа и выделяется около 10 ккал свободной энергии[Мф39].
Ресинтез АТФ Для того чтобы мышечные волокна могли поддерживать сколько-нибудь длительное сокращение, необходимо постоянное восстановление АТФ с такой же скоростью, с какой он расщепляется (расходуется). Энергия для восстановления АТФ образуется за счет питательных («горючих») веществ — углеводов, жиров, белков. При распаде (полном или частичном) этих веществ часть энергии, запасенная в их химических связях, освобождается. Эта свободная энергия обеспечивает связывание АДФ и фосфата с образованием АТФ. Ресинтез АТФ в мышце может происходить двумя основными путями — анаэробным (без участия кислорода) и аэробным (при участии кислорода). Если энергетическое обеспечение работы осуществляется в основном анаэробным путем, такая работа называется анаэробной, а если преимущественно аэробным путем — аэробной работой.
Для образования и использования АТФ в качестве непосредственного источника энергии в сокращающейся мышце могут действовать три химические (энергетические) системы: 1) фосфагенная (АТФ-КФ-система) 2) гликолитическая (лактацидная) 3) окислительная (кислородная)
Фосфагенная и гликолитическая работают по анаэробному пути, окислительная — по аэробному.
Системы отличаются друг от друга по энергетической ёмкости и мощности[Мф40]. Энергетическая ёмкость системы — это максимальное количество образуемой энергии, или максимальное количество АТФ, которое может_ресинтезироваться за счет_энергии рассматриваемой системы (табл. 210210903). Энергетическая мощность системы — это максимальное количество энергии, выделяющейся в единицу времени, или максимальное количество АТФ, образующейся в единицу времени за счет энергии рассматриваемой системы (табл. 210210903). Емкость энергетической системы ограничивает максимальный объем работы, который может быть выполнен за счет данной системы. Мощность системы лимитирует предельную интенсивность работы, выполняемой за счет энергии данной системы.
Таблица 210210903 Максимальная мощность и ёмкость трех энергетических систем (примерные величины из расчета на 20 кг активной мышечной массы*)
* У «стандартного» мужчины весом 70 кг мышечная масса составляет около 40 % веса тела, т.е. примерно 30 кг. При наиболее типичных видах мышечной деятельности (локомоциях) участвует не более 2/3 всей мышечной массы. Общая масса сокращающихся мышц — активная мышечная масса — не превышает обычно 20 кг.
Степень участия трех систем в энергообеспечении (ресинтезе АТФ) зависит от силы и продолжительности мышечных сокращений, т.е. от мощности и длительности работы, а также от условий работы мышц, особенно от уровня их обеспечения кислородом.
Фосфагенная энергетическая система
К этой группе относятся высокоэнергетические фосфатные соединение АТФ и креатинфосфат (КрФ). Восстановление (ресинтез, регенерация) АТФ осуществляется во время мышечных сокращений почти мгновенно, как только он расщепляется до АДФ. КрФ — первый энергетический резерв мышцы, действующий как немедленный источник восстановления АТФ. Конечными продуктами расщепления КрФ являются креатин и свободный фосфат. Энергия от расщепления КрФ немедленно и непосредственно используется для ресинтеза АТФ. При отщеплении фосфатной группы от КрФ освобождается в среднем 10,5 ккал[V.G.41] [Мф42]. Мощность фосфагенной системы наибольшая по сравнению с другими системами (таблица 210210903). Максимальная скорость энергообразования, выраженная в количестве АТФ, ресинтезируемого в единицу времени за счет активности фосфагенной системы, примерно в 3 раза превышает максимальную мощность гликолитической системы и в 4—10 раз — максимальную мощность кислородной системы. Поэтому фосфагенная система играет решающую роль в энергообеспечении работ предельной мощности (спринтерский бег, кратковременные мышечные усилия «взрывного характера» — при подъеме штанги, метаниях, прыжках и т. п.), осуществляемых с максимально возможными по силе и скорости (мощности) сокращениями мышц. Емкость фосфагенной системы невелика, так как запасы АТФ и КФ в мышце весьма ограниченны (таблица 210210903). Емкость фосфагенной системы, оцениваемая запасом АТФ в 20 кг мышц и количеством АТФ, которое может ресинтезироваться за счет КрФ, содержащегося в этой же мышечной массе, составляет всего около 0,5 М АТФ, т.е. около 5 ккал. Поэтому работа с предельной мощностью, которая обеспечивается почти исключительно анаэробной фосфагенной системой, может продолжаться с энергетической точки зрения не более нескольких секунд. Например, при спринтерском беге энергетический запрос работающих мышц может достигать около 1 ккал/с. Поскольку максимальное количество энергии, которое может обеспечить фосфагенная система, составляет около 5 ккал, такая работа теоретически может продолжаться за счет фосфагенной системы не более 5 с. Если выполняется работа большей продолжительности, её энергетическое обеспечение не может происходить только за счет наличных запасов мышечных фосфагенов (АТФ и КрФ).
Гликолитическая энергетическая система
В основе деятельности этой системы, обеспечивающей ресинтез АТФ и КФ, лежит цепь химических реакций анаэробного расщепления гликогена (и глюкозы). Совокупность этих реакций называется анаэробный гликогенолиз, или гликолиз (или цикл Эмбден — Мейерхофа). В результате таких реакций образуется молочная кислота — лактацид. Отсюда другое название этой энергетической системы — лактацидная. В результате гликолиза освобождается энергия, которая позволяет ресинтезировать АТФ. Энергетическими субстратами лактацидной мышечной системы служат углеводы, главным образом гликоген, запасенный в мышечных клетках, и в меньшей степени — глюкоза. Молекула гликогена — это цепочка молекул глюкозы. При расщеплении молекулы гликогена от нее последовательно отщепляются молекулы глюкозы (глюкозные единицы). В мышечные клетки поступает также глюкоза из крови. В свою очередь, глюкоза попадает в кровь из печени. Анаэробное расщепление гликогена (гликогенолиз) в мышечных клетках имеет энергетическое преимущество перед гликолизом (анаэробным расщеплением глюкозы), так как каждая глюкозная единица из молекулы мышечного гликогена в процессе гликолиза дает энергию для образования 3 молекул АТФ, а каждая молекула глюкозы — только для 2 молекул АТФ. В результате анаэробного гликогенолиза из каждой глюкозной единицы (глюк. ед.) образуются 2 молекулы молочной кислоты. Химические реакции анаэробного гликогенолиза (гликолиза) происходят исключительно в саркоплазме, где находятся все ферменты, катализирующие гликолиз (гликолитические ферменты). Анаэробный гликогенолиз развертывается практически с начала мышечной работы, но достигает максимальной мощности (наибольшей скорости образования молочной кислоты) лишь через 30—40 с. Поэтому наибольшие концентрации молочной кислоты в мышцах (и в крови) обнаруживаются лишь при работах, которые продолжаются более 20—30 с. При более кратковременной работе энергетическая роль лактацидной системы относительно меньше. Мощность лактацидной системы мышц велика. При работах очень большой мощности, которые могут продолжаться лишь несколько десятков секунд, скорость расходования гликогена в мышцах достигает 10 мМ глюк.ед/кг веса мышцы в минуту или около 0,2 г/мин (рис[V.G.43].). Определение содержания и расходования в мышцах гликогена и других энергетических субсгратов у человека стало возможным благодаря использованию метода игольчатой биопсии — взятию малой порции мышечной ткани с помощью специальной иглы (Д.Бергстрём) до работы и в разные периоды ее выполнения, а также после работы. Таким образом, скорость расщепления гликогена в активной мышечной массе в 20 кг равна 200 мМ глюк. ед/мин. Если расщепление гликогена идет анаэробным путем, то при этом образуется 400 мМ молочной кислоты в минуту. Образование каждой молекулы молочной кислоты из гликогена обеспечивает ресинтез 3 молекул АТФ. Следовательно, при образовании 400 мМ молочной кислоты ресинтезируется 1,2 М АТФ/мин. Такова примерная максимальная мощность лактацидной системы (см. табл. 2). Мощность лактацидной системы в 1,5 раза выше, чем кислородной, но примерно в 3 раза ниже мощности фосфагенной системы (табл. 210210903). Лактацидная система играет решающую роль в энергетическом обеспечении работ очень большой мощности, которые могут продолжаться от 20 с до 1—2 мин и связаны с сильными мышечными сокращениями, требующими большой скорости расщепления и соответственно ресинтеза АТФ. К таким видам мышечной работы относятся, например, бег на дистанциях от 200 до 800 м, плавание на дистанциях от 50 до 200 м и т.д. По мере дальнейшего уменьшения мощности выполняемой работы (увеличения её продолжительности) энергетическая роль лактацидной системы снижается. Лактацидная система функционирует в тех случаях, когда сокращающиеся мышцы испытывают недостаток в снабжении кислородом. Такие условия возникают: при работах очень большой мощности, в самом начале любой работы когда снабжение мышц кислородом отстает от потребности в нем [p], при статическом сокращении мышц даже небольшой силы (превышающей 20% от максимальной).
При статическом сокращении мышц даже небольшой силы (превышающей 20% от максимальной) из-за внутримышечного давления резко ограничивается кровоснабжение, а, следовательно, и обеспечение мышц кислородом. Образующаяся в процессе анаэробного гликогенолиза (гликолиза) молочная кислота медленно диффундирует в кровь. Поэтому по мере образования она скапливается внутри сокращающихся мышечных клеток, что ведет к сдвигу их активной реакции в кислую сторону. По мере накопления молочной кислоты происходит угнетение вплоть до полного торможения активности ключевых гликолитических ферментов мышечных клеток, в частности фосфорилазы и фосфофруктокиназы. В результате снижается скорость гликолиза (образования молочной кислоты) и количество энергии (АТФ), образующейся в единицу времени за счет лактацидной системы. Таким образом, скорость образования (концентрация) в сокращающихся мышцах молочной кислоты регулируется по механизму отрицательной обратной связи — торможением — конечным продуктом реакции: чем больше скорость образования (накопления) молочной кислоты, тем сильнее торможение, замедляющее анаэробный гликогенолиз (гликолиз). Поэтому емкость лактацидной системы в основном лимитируется не содержанием ее энергетических субстратов — углеводов (мышечного гликогена и глюкозы крови), а количеством (концентрацией) конечного продукта деятельности этой системы — молочной кислоты. В этой связи при выполнении, мышечной работы, энергетическое обеспечение которой осуществляется преимущественно лактацидной системой, никогда не происходит резкого истощения гликогена в работающих мышцах, а тем более в печени.
Ёмкость лактацидной системы. Подсчитаем её. Среднее содержание гликогена в мышцах — около 15 г/кг, или около 80 мМ глюк. ед (молекулярный вес глюкозы = 180). При работах с максимальной для лактацидной системы нагрузкой в результате анаэробного гликогенолиза в рабочих мышцах расходуется, по-видимому, не более 20 мМ глюк. ед/кг. Таким образом, во всей активной мышечной массе (20 кг) расходуется в целом примерно 400 мМ, или 0,4 М глюк. ед. При анаэробном глюкогенолизе 1 М глюк. ед. дает энергию, достаточную для образования 3 М АТФ. Следовательно, при анаэробном расщеплении 0,4 М глюк. ед. возможен ресинтез 1,2 М АТФ, что эквивалентно общей энергопродукции за счет анаэробного расщепления мышечного гликогена в 12000 ккал. Эти величины и характеризуют максимальную энергетическую емкость лактацидной системы (см. табл. 210210903).
Другой способ подсчета емкости лактацидной системы — по максимальному количеству образующейся в мышцах молочной кислоты. Максимальная концентрация молочной кислоты в работающих мышцах достигает у нетренированного человека 250 мг/100 г., или 2,5 г/кг. В этом случае во всей массе активных мышц (20 кг) общее содержание молочной кислоты составляет 50 г, а при учете ее молекулярного веса (90) — около 0,6 М. При анаэробном расщеплении одной глюкозной единицы гликогена освобождается энергия, достаточная для ресинтеза 3 молекул АТФ. При этом образуются 2 молекулы молочной кислоты. Следовательно, при образовании 0,6 М молочной кислоты освобождается энергия, достаточная для ресинтеза — 0,9 М АТФ (около 9 ккал). В действительности же при работе общее количество образующейся в мышцах молочной кислоты превышает рассчитанное по ее максимальной концентрации, так как молочная кислота непрерывно диффундирует из мышечных клеток в кровь. Следовательно, и емкость лактацидной системы должна превышать те величины, которые рассчитываются по максимальной концентрации лактата. Энергетическая емкость лактацидной системы примерно в 2,5 раза больше, чем фосфагенной системы (табл. 210210903).
Кислородная (окислительная) энергетическая система При непрерывном поступлении кислорода в митохондрии мышечных клеток действует кислородная система энергопродукции (ресинтеза АТФ). При работах аэробного характера с повышением интенсивности (мощности) выполняемой нагрузки увеличивается количество O2, потребляемого мышцами (и всем организмом в целом) в единицу времени. Поскольку между скоростью потребления O2 (л/мин) и мощностью работы (Вт) аэробного характера существует прямо пропорциональная (линейная) зависимость, интенсивность аэробной работы можно характеризовать скоростью потребления кислорода (рис[V.G.44].). При определенной, индивидуальной для каждого человека нагрузке достигается максимально возможная для него скорость потребления O2 — максимальное потребление кислорода (МПК). Для физиологической оценки относительной мощности выполняемой человеком аэробной мышечной работы её оценивают по относительной скорости потребления O2, т.е. выраженному в процентах отношению скорости потребления O2 (л/мин) при выполнении данной работы к МПК (% МПК).
Энергетические субстраты. Для энергетического обеспечения мышечной работы кислородная система может использовать в качестве субстратов окисления («горючего») все основные питательные вещества — углеводы (гликоген и глюкозу), жиры (жирные кислоты) и белки (аминокислоты). Вклад белков (аминокислот) в общую аэробную энергопродукцию мышц весьма мал и потому может практически не учитываться. До тех пор, пока углеводы и жиры доступны мышце, именно они используются как субстраты аэробной (окислительной) энергопродукции для поддержания мышечных сокращений.
Соотношение между окисляемыми углеводами и жирами определяется относительной мощностью аэробной работы (% МПК): чем больше относительная мощность выполняемой работы, тем относительно выше энергетический вклад окисляемых углеводов и соответственно меньше вклад окисляемых жиров в общую энергопродукцию работающих мышц (рис[V.G.45].). Во время выполнения легкой работы при потреблении O2 до 50 % МПК (с предельной продолжительностью до нескольких часов) большая часть энергии для сокращающихся мышц образуется за счет окисления жиров. Во время более тяжелой работы (при потреблении O2 более 60 % МПК) значительную часть энергопродукции обеспечивают углеводы. При работах, близких к МПК, подавляющая часть аэробной энергопродукции идет за счет окисления углеводов. Таким образом, при работе очень большой мощности основными энергетическими субстратами в работающих мышцах служат углеводы. Они расщепляются главным образом аэробно (окисляются) при работе продолжительностью до нескольких десятков минут и в значительной мере анаэробно (гликолитически) при менее продолжительной работе. Аэробное расщепление углеводов (гликогена и глюкозы) вплоть до стадии образования пировиноградной кислоты идет по тому же пути, что и при анаэробном гликолизе. В последнем случае из-за недостатка O2 (или уменьшения отношения НАД / НАДН+) пировиноградная кислота превращается (восстанавливается) в молочную кислоту при участии фермента лактатдегидрогеназы (ЛДГ). В аэробных условиях пировиноградная кислота не восстанавливается до молочной кислоты, а окисляется. При этом образуются конечные продукты окисления — CO2 и H2O. Суммарное уравнение для аэробного расщепления (окисления) глюкозы (C6H12O6) выглядит следующим образом:
Из этого уравнения следует, что при полном аэробном расщеплении одной молекулы глюкозы образуются 38 молекул АТФ, из которых 2 молекулы АТФ образуются на гликолитическом этапе расщепления глюкозы до пировиноградной кислоты, а 36 молекул АТФ — за счет кислородной системы. На этапах анаэробного расщепления из одной глюкозной единицы мышечного гликогена образуются 3 молекулы АТФ. Поэтому при полном окислении одной глюкозной единицы гликогена образуются 39 молекул АТФ (3 + 36). Следовательно, аэробное расщепление глюкозы дает в 19 раз больше продукции АТФ, чем анаэробный гликолиз (38 молекул АТФ против 2 молекул АТФ). Окисление глюкозной единицы из мышечного гликогена обеспечивает образование в 13 раз большего числа молекул АТФ, чем при анаэробном гликогенолизе (39 молекул АТФ против 3 молекул АТФ). Таким образом, с точки зрения расходования горючего (углеводов) кислородная система значительно более эффективна, чем анаэробная лактацидная: при одинаковом расходовании углеводов за счет их окислительного расщепления ресинтезируется во много раз больше АТФ, чем при анаэробном гликогенолизе (гликолизе). Приведенное выше уравнение позволяет также вычислить кислородный эквивалент АТФ при окислении углеводов, т.е. количество O2, необходимое для окислительного ресинтеза 1 моля АТФ.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-14; просмотров: 858; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.12.36.30 (0.126 с.) |











 Рис. 210201621. Микрофотография участка мышечного волокна. МФ – миофибрилла, А – анизотропный диск (А-диск), I – изотропный диск (I-диск), H – H‑полоска, S – S‑зона, Z - Z-мембрана, M – M‑мембрана.
Рис. 210201621. Микрофотография участка мышечного волокна. МФ – миофибрилла, А – анизотропный диск (А-диск), I – изотропный диск (I-диск), H – H‑полоска, S – S‑зона, Z - Z-мембрана, M – M‑мембрана.
 Рис. 210211130. Схема саркомера.
Рис. 210211130. Схема саркомера.

 Рис. 709240110. Упрощённая схема саркомера.
Рис. 709240110. Упрощённая схема саркомера.


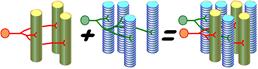 Рис. 709261947. Двигательные единицы в скелетной мышце.
Рис. 709261947. Двигательные единицы в скелетной мышце.
 Рис. 709240129. Микрофотография миофибриллы при сокращении. МФ – миофибрилла, Z - Z-мембрана, M – M‑мембрана.
Рис. 709240129. Микрофотография миофибриллы при сокращении. МФ – миофибрилла, Z - Z-мембрана, M – M‑мембрана.
 Рис. 709240133. Изменение саркомера при сокращении. Объяснение в тексте.
Рис. 709240133. Изменение саркомера при сокращении. Объяснение в тексте.
 Рис. 709270838. Изменение саркомера при сокращении (упрощённая схема). Объяснение в тексте.
Рис. 709270838. Изменение саркомера при сокращении (упрощённая схема). Объяснение в тексте.

 Рис. 709270834 показывает цикл миозиновых мостиков без деталей и может быть использован студентом при ответе.
Рис. 709270834 показывает цикл миозиновых мостиков без деталей и может быть использован студентом при ответе.
 Рис. 709270834. Рабочий цикл миозиновых мостиков поперечнополосатого миоцита.
Рис. 709270834. Рабочий цикл миозиновых мостиков поперечнополосатого миоцита.
 1. Головка миозина, несущая продукты гидролиза АТФ (АДФ + фосфат) соединяется с миозинсвязывающим участкам актиновой нити.
2. АДФ и фосфат покидают миозиновую головку. Головка миозина изменяет конформацию за счёт шарнирного участка в области шейки миозина. Происходит гребковое движение, продвигающее актиновую нить к центру саркомера (рабочий ход).
3. Головка миозина связывается с молекулой АТФ, что приводит к отделению миозина от актина.
4. Гидролиз АТФ восстанавливает конформацию молекулы миозина, и она оказывается готовой вступить в новый цикл.
1. Головка миозина, несущая продукты гидролиза АТФ (АДФ + фосфат) соединяется с миозинсвязывающим участкам актиновой нити.
2. АДФ и фосфат покидают миозиновую головку. Головка миозина изменяет конформацию за счёт шарнирного участка в области шейки миозина. Происходит гребковое движение, продвигающее актиновую нить к центру саркомера (рабочий ход).
3. Головка миозина связывается с молекулой АТФ, что приводит к отделению миозина от актина.
4. Гидролиз АТФ восстанавливает конформацию молекулы миозина, и она оказывается готовой вступить в новый цикл.






