Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Механізми транспорту речовин через мембрану
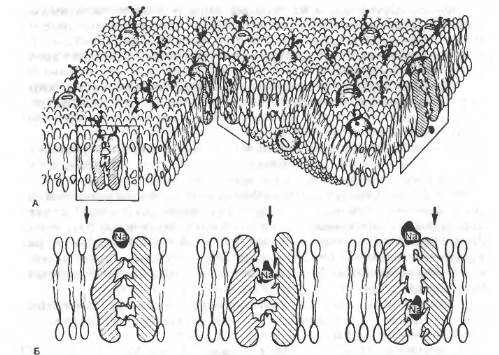
Усі механізми транспорту речовин через мембрани клітин поділяються на: І. Пасивні (не потребують затрат енергії). 1. Проста дифузія. 2. Полегшена дифузія. 3. Осмос. ІІ. Активні (потребують затрати енергії, тобто АТФ). 4. Первинний активний транспорт. 5. Вторинний активний транспорт. 6. Піноцитоз. Найпоширенішим і найпростішим способом переміщення речовин через мембрани клітин є дифузія. Завдяки дифузії більшість молекул у водних розчинах (а у людини від 45% до 80% води в організмі) вільно проникає через мембрани за градієнтом концентрації (від середовища з більшою концентрацією даної речовини до середовища з меншою). При цьому концентрація цих речовин по обидва боки мембрани прагне до зрівноваження. Мембрани дифузії не перешкоджають, тому така дифузія називається простою (1). Таким чином через ліпідні мембрани клітин проникає (дифундує) вода, молекули кисню і вуглекислого газу, жиророзчинні речовини, етанол, сечовина. Цукор, заряджені молекули, в т.ч. неорганічні іони, білки, амінокислоти важко проходять через мембрани. Для неорганічних іонів, наприклад, у клітинній оболонці існують пори, так звані канали, утворені зануреними у мембрани транспортними білками. Всередині таких білків існує заповнений водою отвір, діаметром менше 1 нм, через який може дифундувати мала молекула, яка також рухається за градієнтом концентрації. Мембранні канали специфічні, тобто вибірково пропускають молекули. Наприклад, калієвий канал проникний в основному для іонів К+.
Рис.1. А. Біліпідна мембрана клітини з вбудованими каналами. Б. Натрієві канали.
Глюкоза та більшість амінокислот проникають у клітину шляхом полегшеної дифузії (2) – за допомогою білків-носіїв. Ще одним видом пасивного транспорту є осмос (3), який має значення для напівпроникних мембран, тобто мембран, проникних лише для води і непроникних для інших іонів. При цьому молекули води рухаються у бік більшої концентрації речовин (наприклад, Na+, глюкози). Так як вода є розчинником, то поступово усувається різниця концентрації розчиненої у ній речовини. Але у процесі розвитку механізму збудливості іноді виникають моменти, коли для того, щоб досягнути кількісної стабільності іонів, необхідно їх переносити через мембрану всупереч електричному чи концентраційному градієнту, тобто у бік їх більшої концентрації. Такий транспорт називається активним (ІІ). Він здійснюється за допомогою іонних помп (насосів) та енергії АТФ. Практично у кожній клітині існують Na/K помпи, які викачують іони натрію з клітини і одночасно закачують у неї іони калію. Т.ч. забезпечується низька внутрішньоклітинна концентрація Na+ і висока концентрація К+. Різні клітини витрачають на роботу Na/K помпи від 30 до 70% своєї енергії. При цьому АТФ розщеплюється до АДФ і фосфату за допомогою ферменту АТФ-ази, розміщеного на мембрані клітини: 1 молекула АТФ забезпечує клітині вихід 3 іонів Na та вхід 2 іонів К.
Крім Na/K помпи існує кальцієва, яка відкачує Са2+ з клітини. Кальцієвих помп особливо багато в ендоплазматичній сітці м’язових клітин, яка накопичує іони кальцію. Якщо через помпи проходить один вид іонів, то такий транспорт називається первинним активним (4). Існуєтакож вторинний активний транспорт (5), який забезпечує рух двох видів молекул чи іонів або в одному напрямку (наприклад, разом з іоном натрію у клітину можуть увійти глюкоза чи амінокислоти) - симпорт (5а),або у протилежних напрямках (наприклад, у клітину заходить натрій, а виходить кальцій) – антипорт (5б). Для білків і холестеролу, які є великими молекулами, відсутні транспортні канали, тому вони проходять шляхом піноцитозу (6) у вигляді пухирців (везікул). Такі пухирці або зливаються з мембраною клітини, даючи можливість вмісту клітини вийти з неї (екзоцитоз) або осідають на рецепторах мембран, ділянки яких вгинаються, заглиблюються, утворюючи ямку, яка згодом змикається, формуючи внутрішньоклітинний пухирець (ендоцитоз).
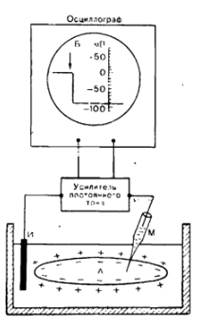
Функціонування збудливих тканин пов’язане з рухом заряджених частинок, тобто іонів Na, K, Са тощо. Так як іони, які рухаються через мембрану, несуть на собі заряд, цей заряд («+» або «-») накопичується з одного чи іншого боку мембрани, створюючи різницю потенціалів між зовнішнім та внутрішнім боком мембрани – мембранний потенціал. Мембранний потенціал можна виміряти за допомогою осцилографа з підсилювачем, якщо один електрод встановити на поверхні клітини, а інший ввести у клітину (всередині минулого століття винайшли такий мікроелектрод). У момент введення мікроелектроду стрілка осцилографа відхилиться, що засвідчує наявність електричного потенціалу на мембрані. Подальше просування мікроелектроду всередині клітини не впливає на стрілку осцилографа. Це підтверджує те, що потенціал дійсно локалізується на клітинній мембрані.
Дифузія різних іонів через мембранні канали повинна була б привести до усунення різниці концентрацій між поза- та внутрішньоклітинним середовищем, але цього не спостерігається, так як різниця в концентрації іонів між клітиною та позаклітинним середовищем дуже велика, про що згадувалося вище. Коли клітина збудливої тканини знаходиться у спокої (тобто не збуджена), відкриті, в основному, калієві канали і незначна кількість натрієвих. При цьому за градієнтом концентрації (від більшої концентрації до меншої) іон К+ виходить з клітини та виносить 1 позитивний заряд. Отже, внутрішньоклітинне середовище стає більш негативно зарядженим, а на мембрані виникає різниця потенціалів, яку назвали мембранним потенціалом спокою. Терміном «мембранний потенціал» (МП) (потенціал спокою, ПС) називають різницю потенціалів, яка існує між цитоплазмою (внутрішньою стороною мембрани) та оточуючим клітину зовнішнім розчином. Коли клітина знаходиться у стані фізіологічного спокою, її внутрішній потенціал є негативним по відношенню до зовнішнього Внутрішньоклітинний негативний заряд перешкоджає виходу з клітин нових іонів К, а ті іони, які зможуть вийти, ще більше підвищать заряд на мембрані. Потік іонів К припиняється, коли дія електричного поля компенсує дифузійний тиск внаслідок різниці потенціалів. При цьому іони продовжують проходити через мембрану, але у рівних кількостях в обох напрямках. Було виявлено, що всі клітини мають мембранний потенціал (МП). Наприклад, у м’язових клітинах ссавців його рівень становить близько -90 мВ. Інші клітини можуть мати МП в межах від -40 до -120 мВ. В основному, даний рівень МПС забезпечується виходом іонів К+ за межі клітини, але дещо (на 10 мВ) його величина зменшується завдяки незначному русі іонів Na+ у клітину. Здатність різних заряджених частинок проникати через клітини мембрани різна і залежить від: виду іона (його проникності), товщини мембрани, температури середовища, наявності деяких речовин (наприклад, динітрофенолу, який вичерпує запаси енергії), рівня насиченості іонів у середовищах тощо. За однакових умов для більшості клітинних мембран проникність для іонів К (РК) приблизно в 30 разів вища за РNa. РCl сильно коливається: для більшості мембран РCl мала по відношенню до РК, а в скелетних м’язах значно вища, ніж РК. Мембранний потенціал регулюється рівнем насиченості іонів. При зменшенні внутрішньоклітинної та підвищенні позаклітинної концентрації К+ потенціал мембрани зменшується або навіть змінюється його знак (потенціал стає позитивним, якщо у зовнішньому розчині концентрація К+ була вищою, ніж у внутрішньому). Іони Cl- у нервових волокнах не відіграють істотної ролі, оскільки проникність для них мембрани у стані спокою відносно мала. Але у скелетних м’язових волокнах проникність для Cl- достатня, щоб збільшити їх потенціал спокою.
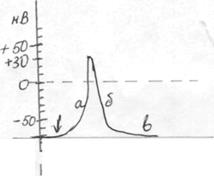
Регулювати проходження іонів через канали можна також за допомогою специфічних хімічних речовин – блокаторів. Так, блокатором натрієвих каналів є тетродотоксин, який синтезується у деяких риб; калієвих – тетраетиламоній, амінопіридини, кальцієвих – Mn2+, верапаміл, ніфедепін. Блокатори використовують у клініці для пригнічення підвищеної активності м’язів тощо. Регулювати роботу помп також можна за допомогою речовин, які впливають на окремі ланки механізму перекачування іонів через помпу. Наприклад, серцевий глікозид уабаїн (основа препарату строфантину) гальмує активність АТФ-ази (фермент, що розщеплює АТФ, який необхідний для роботи помпи), внаслідок чого порушується робота помпи, клітина губить К+ та збагачується Na+. А потужний пригнічувач (інгібітор) дихальних ферментів ціанід гальмує утворення АТФ. Без своєчасного введення АТФ він може зупинити роботу помп і спричинити навіть смерть. Потенціал дії Основна функція нервових клітин в організмі полягає в отриманні інформації, передачі її до інших відділів нервової системи, у регуляції дії інших клітин. Це забезпечується виникненням у них явища збудження, яке реалізується через утворення потенціалу дії (ПД). ПД -це швидкі коливання мембранного потенціалу. В його основі - зміна проникності мембран для іонів. Амплітуда і характер змін ПД мало залежать від сили подразника. Важливо, щоб ця сила досягнула деякої порогової величини. Наявність порогу і залежність амплітуди ПД від сили стимулу отримали назву закону «все або нічого». Причиною виникнення ПД є подразнення рецепторів, або збудження нервових клітин. Для реєстрації ПД використовують поза- і внутрішньоклітинні електроди. Поза-клітинні електроди дозволяють виявити, що поверхня збудженої ділянки на дуже короткий час стає негативно зарядженою порівняно з сусідньою ділянкою, яка знаходиться у стані спокою. Внутрішньоклітинні електроди дозволяють кількісно охарактеризувати зміни мембранного потенціалу під час висхідної та низхідної фаз ПД. За допомогою цього методу було встановлено, що потенціал дії складається з наступних фаз: 1. Фаза деполяризації (висхідна). У ній виникає різниця потенціалів зворотного знаку: внутрішній вміст клітини стає зарядженим позитивно порівняно з зовнішнім середовищем. 2. Фаза реполяризації (низхідна). МП повертається до вихідного Обидві фази утворюють пік потенціалу дії і залежать від температури середовища. 3. Слідові потенціали, якіспостерігаються після піку і можуть бути у вигляді слідової деполяризації (у скелетних м'язових волокнах) або слідової гіперполяри-зації (у нервових волокнах).
: ПД аксону кальмара: а) фаза деполяризації; а) фаза деполяризації: б) фаза реполяризації; б) фаза реполяризації; в) фаза слідової деполяризації; в) фаза слідової гіперполяризації. ↓- момент нанесення подразника.
|
|||||||
|
|
Последнее изменение этой страницы: 2016-08-14; просмотров: 1215; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.161.116 (0.016 с.) |