Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Эндокринной систем и органов чувств в филогенезеСтр 1 из 12Следующая ⇒
Эндокринной систем и органов чувств в филогенезе Беспозвоночных и позвоночных. Учебно-методическое пособие для студентов 1-го курса СтГМА
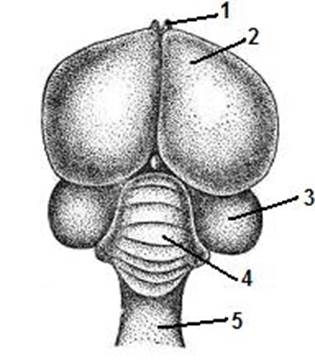
Ставрополь 2008 Рис. 4. Схема формирования нервной трубки зародыша цыпленка (по А.Г. Кнорре). А – стадия нервной пластинки. Б – стадия нервной трубки. В – обособление нервной трубки и ганглиозной пластинки от эктодермы; Нервный желобок; 2 – нервные валики; 3 – кожная эктодерма; 4 – хорда; 5 – мезодерма; 6 – ганглиозная пластинка; 7 – нервная трубка; 8 – мезенхима; 9 – невроцель.
Образование у позвоночных головного мозга (кефализация, или цефализация) было связано с усилением у них двигательной активности и необходимостью постоянного анализа информации, поступающей из органов чувств. Полагают, что передний мозг сформировался в ходе развития динамических координаций с органом обоняния, средний - с органом зрения, а задний – со статокинетическим анализатором. Внутри головного и спинного мозга на базе невроцеля образуется полость, заполненная спинномозговой жидкостью. В головном мозге эта полость представлена сообщающимися между собой цистернами - мозговыми желудочками. Спинномозговая жидкость, заполняющая их, образуется в сосудистых сплетениях за счет фильтрации плазмы крови.
Рис. 5. Развитие зачатка головного мозга позвоночных. В мозговых желудочках различают дно (основание) и крышу (мантия). Крыша располагается над желудочками, а дно - под ними. В веществе головного мозга нейроны распределяются не диффузно, а в форме скоплений, образуя серое вещество, а их отростки - белое вещество. Слой серого вещества в крыше любого отдела мозга называется корой, а отдельные скопления его в толще белого вещества - ядрами.
Сравнительно-анатомический обзор головного мозга позвоночных. Изучение сравнительно-анатомических особенностей органов и систем органов имеет не только важное общебиологическое значение для решения вопросов филогенетического родства и общности происхождения тех или иных групп организмов. Эта проблема является одним из важнейших звеньев в медицинском образовании, поскольку вся морфологическая и функциональная сущность человека является результатом длительной эволюции его животных предков и, прежде всего, позвоночных. И только зная пути и закономерности филогенетических преобразований тех или иных органов, можно правильно оценить их резервные возможности в восстановительных процессах и правильно понять предпосылки и механизмы развития тех или иных аномалий.
Рыбы. У представителей этого класса в строении головного мозга наблюдаются вариации, но, тем не менее, можно выделить общие характерные для них черты. Их головной мозг имеет относительно примитивное строение и в целом небольшие размеры. Передний мозг, или конечный, у большинства рыб состоит из одного полушария (у некоторых акул, ведущих придонный образ жизни, их два) и одного желудочка. Крыша не содержит нервных элементов и образована эпителием и лишь у акуловых нервные клетки поднимаются от основания мозга на бока и отчасти на крышу. Дно мозга представлено двумя скоплениями нейронов - это полосатые тела (corpora striata). Впереди от мозга располагаются две обонятельные доли (луковицы), связанные обонятельными нервами с органом обоняния, расположенным в ноздрях. У низших позвоночных передний мозг представляет собой отдел нервной системы, обслуживающий только обонятельный анализатор. Он является высшим обонятельным центром. Промежуточный мозг состоитиз эпиталамуса, таламуса и гипоталамуса, которые характерны для всех позвоночных, хотя степень их выраженности варьирует. Особую роль в эволюции промежуточного мозга играет таламус,в котором выделяют вентральную и дорсальную части. В дальнейшем у позвоночных в ходе эволюции размеры вентральной части таламуса уменьшаются, а дорсальной увеличиваются. Для низших позвоночных характерно превалирование вентрального таламуса. Здесь расположены ядра, которые выполняют роль интегратора между средним мозгом и обонятельной системой переднего мозга, кроме того, у низших позвоночных таламус является одним из главных моторных центров. Под вентральным таламусом располагается гипоталамус. Снизу он образует полый стебелек - воронку, которая переходит в нейрогипофиз, соединенный с аденогипофизом. Гипоталамус играет основную роль в гормональной регуляции организма.
Эпиталамус расположен в дорсальной части промежуточного мозга. Он не содержит нейронов и связан с эпифизом. Эпиталамус вместе с эпифизом составляет систему нейрогормональной регуляции суточной и сезонной активности животных.
Ромбовидная ямка. Средний мозг у рыб относительно крупный. В нем различают дорсальную часть - крышу (текум), имеющую вид двухолмия, и вентральную часть, которая называется тегментом и является продолжением двигательных центров ствола мозга. Средний мозг сложился как первичный зрительный и сейсмосенсорный центр. В нем сосредоточены зрительные и слуховые центры. Кроме того, он является высшим интегративным и координирующим центром головного мозга, приближаясь по своему значению к большим полушариям переднего мозга высших позвоночных. Такой тип мозга, где высшим интегративным центром является средний мозг, называется ихтиопсидным. Мозжечок образуется из заднего мозгового пузыря и закладывается в виде складки. Его размеры и форма значительно варьируют. У большинства рыб он состоит из средней части - тела мозжечка и из боковых ушек – аурикул. Для костных рыб характерно переднее разрастание - заслонка. Последняя у некоторых видов принимает столь большие размеры, что может скрывать часть переднего мозга. У акул и костных рыб мозжечок имеет складчатую поверхность, за счет чего его площадь может достигать значительных размеров. Посредством восходящих и нисходящих нервных волокон мозжечок соединяется со средним, продолговатым и спинным мозгом. Его основная функция - это регуляция координации движений, в связи с чем у рыб с высокой двигательной активностью он крупный и может составлять до 15 % от всей массы головного мозга. Продолговатый мозг является продолжением спинного мозга и в целом повторяет его строение. Границей между продолговатым и спинным мозгом считают то место, где центральный канал спинного мозга на поперечном сечении принимает вид круга. При этом полость центрального канала расширяется, образуя желудочек. Боковые стенки последнего сильно разрастаются в стороны, а крышу образует эпителиальная пластинка, в которой располагается сосудистое сплетение с многочисленными складками, обращенными в полость желудочка. В боковых стенках находятся нервные волокна, обеспечивающие иннервацию висцерального аппарата, органов боковой линии и слуха. В дорсальных участках боковых стенок находятся ядра серого вещества, в которых происходит переключение нервных импульсов, поступающих по восходящим проводящим путям из спинного мозга в мозжечок, средний мозг и к нейронам полосатых тел переднего мозга. Помимо этого, здесь также происходит переключение нервных импульсов на нисходящие проводящие пути, связывающие головной мозг с мотонейронами спинного мозга. Рефлекторная деятельность продолговатого мозга очень разнообразна. В нем находятся: дыхательный центр, центр регуляции сердечно-сосудистой деятельности, через ядра блуждающего нерва осуществляется регуляция органов пищеварения, и других органов.
От ствола мозга (средний, продолговатый мозг и варолиев мост) у рыб отходят 10 пар черепно-мозговых нервов.
Земноводные (амфибии). Передний мозг амфибий образует два полушария, внутри которых располагаются латеральные желудочки с сосудистыми сплетениями. Спереди от переднего мозга лежат крупные обонятельные луковицы. Они слабо отграничены от полушарий и у бесхвостых амфибий срастаются между собой по средней линии. Поступающие из обонятельных луковиц сигналы анализируются в переднем мозге, который по существу является высшим обонятельным центром. Крыша переднего мозга образована первичным мозговым сводом – архипаллиумом. В нем находятся нервные волокна (белое вещество), а в глубине, под ними, лежат нервные клетки. На дне переднего мозга имеются скопления нейронов - полосатые тела. Сразу за полушариями переднего мозга располагается промежуточный мозг с хорошо развитыми верхним мозговым придатком - эпифизом и нижним мозговым придатком - гипофизом. В общих чертах промежуточный мозг сходен с таковым у рыб. Наиболее крупным отделом головного мозга у амфибий является средний мозг. Он имеет вид двух полусфер, покрытых корой. В его состав входит зрительный тракт как продолжение зрительных нервов и здесь происходит интеграция зрительного анализатора с другими сенсорными путями и формируется центр, выполняющий сложные ассоциативные функции. Т.о., средний мозг служит ведущим отделом центральной нервной системы, где происходит анализ получаемой информации и вырабатываются ответные импульсы, т.е. у амфибий, как и у рыб, имеется ихтиопсидный тип мозга.
Спинной мозг. Мозжечок у большинства хвостатых и бесхвостых амфибий небольших размеров и имеет вид поперечного валика у переднего края ромбовидной ямки продолговатого мозга. Слабое развитие мозжечка отражает несложную моторную координацию земноводных. Большую часть мозжечка составляет срединная часть (тело мозжечка), где происходит интеграция сигналов от мышечных рецепторов и вестибулярной системы.
У амфибий, как и у рыб, мозжечковые нервные волокна соединены со средним мозгом, стволом и спинным мозгом. Вестибулярно-мозжечковые связи определяют способность животных координировать движения тела. Продолговатый мозг в основных чертах сходен с продолговатым мозгом рыб. От ствола мозга отходит 10 пар черепно-мозговых нервов. Пресмыкающиеся (рептилии). Пресмыкающиеся – это настоящие сухопутные животные, которые могут жить, размножаться и развиваться вдали от водоемов. Они относятся к высшим позвоночным. Их нервная система в связи с подвижным и сложным образом жизни развита лучше, чем у амфибий.
Птицы. Птицы являются высокоспециализированной группой позвоночных, приспособившихся к полету. Будучи физиологически близки к рептилиям, птицы имеют головной мозг во многом сходный с таковым у рептилий, хотя и отличаются рядом прогрессивных черт, что обусловило более сложное и разнообразное поведение. У них головной мозг довольно крупный. Если у рептилий его масса примерно равна массе спинного мозга, то у птиц он всегда больше. Мозговые изгибы резко выражены. Укрупнение головного мозга обусловлено в первую очередь развитием переднего мозга, который по существу прикрывает все отделы головного мозга за исключением мозжечка. Благодаря этому средний мозг не виден сверху, хотя и хорошо развит. Крыша переднего мозга остается слабо развитой. Кора не только не получает дальнейшего развития, но в полушариях исчезает латеральный островок коры, а медиальный сохраняет значение высшего обонятельного центра. Увеличение размеров переднего мозга происходит за счет дна, где располагаются крупные полосатые тела, являющиеся ведущим отделом головного мозга. То есть у птиц сохраняется зауропсидный тип головного мозга. Обонятельные доли очень малы, что связано со слабым развитием обоняния, и тесно примыкают к переднему мозгу. Промежуточный мозг небольшой. У большей части птиц обычно сохраняется только эпифиз, а парапинеальный орган исчезает в позднем эмбриональном периоде. Информация о длине светового дня поступает не от парапинеального органа, а непосредственно от зрительной системы. В таламусе наиболее развит дорсальный отдел, который является переключательным центром афферентных связей с передним мозгом. Он содержит комплекс ядер на пути нисходящих волокон из двигательных центров переднего мозга. Под ним расположен гипоталамус, связанный с гипофизом, который у птиц хорошо развит. Гипоталамус играет основную роль в гормональной регуляции организма, поддержании гомеостаза, полового и пищевого поведения. Средний мозг состоит из двух крупных бугров. В нем сосредоточены высшие центры обработки зрительной и слуховой информации, а так же центры регуляции видоспецифических наследуемых форм скоординированных реакций, которые являются основой жизнедеятельности. Имеются также сенсорные ядра, выполняющие ассоциативную функцию, направляя сигналы в промежуточный и передний мозг. В то же время происходит снижение количества нисходящих связей между крышей среднего мозга и двигательными центрами за счет появления у птиц стриарно-ретикулярных относительно автономных связей между полосатыми телами переднего мозга и ретикулярной формацией ствола мозга.
Мозжечок крупный и его строение усложняется. Спереди он соприкасается с задними краями полушарий переднего мозга, а сзади прикрывает значительную часть продолговатого мозга. В мозжечке различают среднюю часть (червь) и боковые выступы. Его червь испещрен характерными поперечными бороздками. Относительно сложное строение мозжечка обусловлено сложными движениями, требующими высокой координации во время полета. Продолговатый мозг относительно небольшой, его нижняя сторона еще в большей степени, чем у рептилий, образует изгиб вниз, а в области промежуточного мозга имеется изгиб вверх. Черепно-мозговые нервы птиц представлены 12 парами.
Млекопитающие. Млекопитающие – наиболее высокоорганизованный класс позвоночных животных с высокоразвитой центральной нервной системой. В связи с этим приспособительные реакции млекопитающих на условия среды сложны и весьма совершенны. Передний (конечный) мозг крупный, он значительно превосходит все остальные отделы головного мозга. Его полушария разрастаются во всех направлениях, скрывая промежуточный мозг. Средний мозг виден снаружи только у бесплацентарных и низших плацентарных, а у копытных, хищных, китообразных и приматов он покрыт задней частью больших полушарий. У антропоидов и человека затылочные доли переднего мозга надвинуты и на мозжечок. Если первоначально в ходе эволюции основную массу конечного мозга составляли обонятельные доли, то у млекопитающих развитые обонятельные доли имеют только низшие, а у высших обонятельные доли имеют вид небольших придатков, разделенных на обонятельную луковицу и обонятельный тракт. Увеличение относительных размеров переднего мозга млекопитающих связано, прежде всего, с разрастанием его крыши, а не полосатых тел, как у птиц. Мозговой свод (крыша) образован серым веществом, именуемым корой. Последняя представляет собой комплекс, состоящий из древнего плаща (paleopalium), старого (archipallium) и нового плаща (neopalium). Новый плащ занимает промежуточное положение, располагаясь между старым и древним плащами. Старый плащ, или старая кора, располагается медиально и в прошлом его называли гиппокампом или аммоновым рогом. Древний плащ, или древняя кора, занимает латеральное положение.
Новый плащ обычно называют неокортексом (новая кора) и именно из него в основном и состоят полушария переднего мозга. При этом поверхность полушарий может быть гладкой (лисэнцефальной) или складчатой (с бороздами и извилинами). Кроме того, независимо от этого в полушариях выделяют от 4 до 5 долей. Принцип разделения переднего мозга на доли основывается на топографии определенных борозд и извилин. Разделение на доли в лисэнцефаьном (гладком) мозге носит условный характер. Обычно выделяют теменные доли, височные, затылочные и лобные, а у высших приматов и человека еще и пятую долю, которая называется островком. Он образуется в эмбриональном периоде за счет разрастания височной доли на вентральную сторону полушарий. Принимая за исходный тип больших полушарий лисэнцфальный мозг, выделяют три варианта развития рисунка борозд: продольный, дугообразный и «приматный тип». В варианте приматного типа борозда в лобных долях направленны рострально, а в височных – вентро-дорсально На расположение борозд и извилин может значительно влиять форма мозга. У большинства млекопитающих мозг вытянут в ростро-каудальном направлении. Однако у многих дельфинов мозг расширен латерально и относительно укорочен в длину. Для характеристики переднего мозга млекопитающих большое значение, кроме борозд и извилин, имеет характер распределения в коре нейронов (цитоархитектоника). Неокортекс млекопитающих имеет шестислойное строение и характеризуется наличием пирамидных клеток, которые отсутствуют в мозге других позвоночных. Особенно крупные пирамидные клетки (клетки Беца) находятся в двигательной зоне коры. Их аксоны передают нервные импульсы двигательным нейронам спинного мозга и мотонейронам двигательных ядер черепно-мозговых нервов. Различные участки коры больших полушарий являются специализированными зонами обработки информации, поступающей от различных органов чувств. Различаются сенсорные и моторные зоны. Последние формируют нисходящие пути нервных волокон к стволу головного мозга и спинномозговым двигательным ядрам. Между чувствительными и двигательными зонами коры располагаются интегративные участки, которые объединяют входы сенсорных и моторных областей коры и предопределяют выполнение специализированных видоспецифических функций. Кроме этого, имеются ассоциативные зоны коры, не связанные с конкретными анализаторами. Они представляют собой надстройку над остальными участками коры, обеспечивая мыслительные процессы и хранение видовой и индивидуальной памяти. Весь комплекс распределенных в коре зон сопряжен с функциональной специализацией полей. При этом морфологические и функциональные границы полей довольно точно совпадают. Критерием выделения того или иного поля является изменение в распределении клеточных элементов в коре или возникновение в ней нового подслоя. Особенности архитектоники тех или иных полей являются морфологическим выражением их функциональной специализации. Причиной изменения цитоархитектоники в полях служит увеличение количества восходящих и нисходящих нервных волокон. Сейчас созданы топологические карты полей для человека и для многих лабораторных животных. Поля коры головного мозга входят в состав определенных долей и при этом сами подразделяются на функциональные зоны, связанные с конкретными органами или их частями и имеют упорядоченное внутреннее строение. В каждом поле или зоне выделяют так называемые модули вертикальной упорядоченности организации коры. Модуль имеет либо вид колонки либо клубочка, в который включаются нейроны, расположенные по всей толще коры. В колонку входит группа из 110 нейронов, расположенных между парой капилляров, проходящих через поперечник коры. На стадии формировании мозга древнейших гоминид областью, куда было направленно действие естественного отбора, явилась кора и, прежде всего, следующие ее отделы: нижнетеменная, нижняя лобная и височно-теменная области. Преимущество выживания получили те индивиды, а затем и те популяции формирующихся людей которые оказались продвинутыми в отношении развития каких-то элементов частей коры (большей площадью полей более разнообразными и лобильными связями, улучшенными условиями кровообращения и т.д.). развитие новых связей и структур в коре давали новые возможности в отношении изготовления орудий труда и сплочения коллектива. В свою очередь новый уровень техники зачатки культуры, искусства через естественный отбор способствовали развитию мозга. К настоящему времени сформировалось представление о специфическом системокомплексе коры переднего мозга человека, включающем нижнетеменную, заднюю верхневисочную и нижнюю лобную доли коры. Этот комплекс связан с высшими функциями – речью, трудовой деятельностью и абстрактным мышлением. В целом он является морфологическим субстратом второй сигнальной системы. Эта система не имеет собственных переферических рецепторов, а использует старые рецепторные аппараты различных органов чувств. Так,например, установлено, что на языке имеется особая часть тактильного аппарата, развитие которого определяет последовательность звукообразования на начальных этапах формирования членораздельной речи ребенка. К подплащевым структурам переднего мозга относят базальные ядра, полосатые тела (древнее, старое и новое) и септальное поле. В различных отделах переднего и промежуточного мозга располагается комплекс морфофункциональных структур, названных лимбической системой. Последняя имеет многочисленные связи с неокортексом и вегетативной нервной системой. Она интегрирует такие функции мозга, как эмоции и память. Удаление части лимбической системы приводит к эмоциональной пассивности животного, а ее стимуляция к гиперактивности. Важнейшей функцией лимбической системы является взаимодействие с механизмами памяти. Краткосрочная память связана с гиппокампом, а долгосрочная – с неокортексом. Через лимбическую систему происходит и извлечение индивидуального опыта животного из неокортекса, и управление моторикой внутренних органов, и гормональная стимуляция животного. При этом чем ниже уровень развития неокортекса, тем больше поведение животного зависит от лимбической системы, что приводит к доминированию эмоционально-гормонального контроля за принятием решений. У млекопитающих нисходящие связи неокортекса с лимбической системой обеспечивают интеграцию самых разнообразных сенсорных сигналов. С появлением первых зачатков коры у рептилий от плащевой комиссуры отделился небольшой пучок нервных волокон, соединяющих левое и правое полушарие. У плацентарных млекопитающих такой пучок волокон развит значительно больше и называется мозолистым телом (corpus collosum). Последнее обеспечивает функцию межполушарных коммуникаций. Промежуточный мозг, как и у других позвоночных, состоит из эпиталамуса, таламуса и гипоталамуса. Развитие неокортекса у млекопитающих привело к резкому увеличению таламуса, и, прежде всего, дорсального. В таламусе содержится около 40 ядер, в которых происходит переключение восходящих путей на последние нейроны, аксоны которых достигают коры больших полушарий, где обрабатывается информация, поступающая от всех сенсорных систем. При этом передние и латеральные ядра обрабатывают и проводят зрительные, слуховые, тактильные, вкусовые и интероцептивные сигналы в соответствующие проекционные зоны коры. Есть мнение, что болевая чувствительность не проецируется в кору полушарий переднего мозга, а ее центральные механизмы находятся в таламусе. Это предположение основывается на том, что раздражение разных областей коры не вызывает боли, в то время как при раздражении таламуса ощущается сильная боль. Часть ядер таламуса являются переключательными, а другая часть ассоциативными (от них идут пути в ассоциативные зоны коры). В медиальной части таламуса находятся ядра, которые при низкочастотной электрической стимуляции вызывают в коре больших полушарий развитие тормозных процессов, приводящих ко сну. Высокочастотная стимуляция этих ядер вызывает частичную активацию корковых механизмов. Таким образом таламокортикальная регулирующая система, контролируя потоки восходящих импульсов, участвует в организации смены сна и бодрствования. Если у низших позвоночных высшие сенсорные и ассоциативные центры находятся в среднем мозге, а дорсальный таламус является скромным интегратором между средним мозгом и обонятельной системой, то у млекопитающих он является важнейшим центром переключения слуховых и соматосенсорных сигналов. При этом соматосенсорная область превратилась в наиболее заметное образование промежуточного мозга и играет огромную роль в координации движений. Следует заметить, что комплекс ядер таламуса формируется как за счет зачатка промежуточного мозга, так и за счет миграции из среднего мозга. Гипоталамус образует развитые латеральные выпячивания и полый стебелек - воронку. Последний в заднем направлении заканчивается нейрогипофизом плотно соединенным с аденогипофизом. Гипоталамус является высшим центром регуляции эндокринных функций организма. Он объединяет эндокринные механизмы регуляции с нервными. Кроме того, он является высшим центром симпатического и парасимпатического отделов вегетативной нервной системы. Эпиталамус служит нейрогуморальным регулятором суточной и сезонной активности, что сочетается с контролем за половым созреванием животных. Средний мозг образует четверохолмие, передние бугры которого связаны со зрительным анализатором, а задние – со слуховым. По соотношению относительных размеров передних и задних бугров можно судить о том, какая из систем, слуховая или зрительная, является превалирующей. Если лучше развиты передние бугры, значит, зрительная афферентация (копытные, многие хищники и приматы), если задние – то слуховая (дельфины, летучие мыши и др.). Тегмент подразделяется на чувствительную и моторную зоны. В моторной зоне располагаются двигательные ядра черепно-мозговых нервов и нисходящие и восходящие спинноцеребральные волокна. В связи с развитием у млекопитающих неокортекса как высшего интегративного центра врожденные реакции среднего мозга позволили коре «не заниматься» примитивными формами видоспецифичных реакций на внешние сигналы, в то время как сложные ассоциативные функции принимают на себя специализированные поля коры. Мозжечок у млекопитающих приобретает наиболее сложное строение. Анатомически в нем можно выделить среднюю часть – червь, расположенные по обе стороны от него полушария и флоккулонодулярные доли. Последние представляют филогенетически древнюю часть – архицеребеллум. Полушария в свою очередь делятся на переднюю и заднюю доли. Передние доли полушарий и задняя часть червя мозжечка представляют филогенетически старый мозжечок – палеоцеребеллум. Филогенетически самая молодая часть мозжечка - неоцеребеллум включает в себя переднюю часть задних долей полушарий мозжечка.
В полушариях мозжечка выделяют верхнюю поверхность, образующую кору мозжечка, и скопления нервных клеток – ядра мозжечка. Кора мозжечка построена по единому принципу и состоит из 3 слоев. Мозжечок связан с другими отделами центральной нервной системы тремя парами ножек, образованных пучками нервных волокон. Задние ножки состоят преимущественно из проприоцептивных волокон, которые приходят из спинного мозга. Средние ножки состоят из волокон, соединяющих мозжечок и кору переднего мозга, а передние ножки сформированы нисходящими волокнами, соединяющими мозжечок и средний мозг. Вестибуломозжечковые связи определяют способность животных координировать движения тела, что является основной функцией архицеребеллума. Кроме этого, у млекопитающих сформировались новые более мощные мозжечковые пути за счет возникновения зубчатого ядра мозжечка. Оно получает волокна от различных участков полушарий мозжечка и передает сигналы в таламус, где происходит интеграция сенсомоторных сигналов с активностью корковых центров переднего мозга. Эволюция мозжечка приводит не только к дублированию его древних связей, но и к формированию новых путей. Так, возникает связь через зубчатое ядро с вентролатеральным ядром таламуса и ретикулярными ядрами ствола мозга позволяют поддерживать мышечный тонус и осуществлять рефлекторные реакции. Связи с вестибулярным центром позволяют осуществлять контроль за положением тела в пространстве, а таламические связи предопределяют тонкие сенсомоторные координации. Все эти процессы осуществляются за счет сложной системы межклеточных взаимодействий на уровне коры мозжечка.
Особенности морфо-функциональных преобразований Нервной системы у хордовых.
Первые признаки вегетативной нервной системы намечаются у ланцетника, у которого из дорсальных корешков нервной трубки отходят нервы, идущие к диффузным сплетениям нейронов во внутренних органах. В последних еще нет специализированных ганглиев. Есть предположение, что это признаки парасимпатической системы. У круглоротых имеются коллатеральные ганглии, которые собирают часть висцеральных волокон, расходящихся от ганглиев к внутренним органам. Однако симпатическая цепочка ганглиев еще отсутствует. У позвоночных значительного развития достигает симпатический отдел нервной системы. Они состоит в основном из нервных узлов расположенных около позвоночного столба и связанных продольными тяжами нервных пучков. Вегетативная нервная система хорошо развита у хрящевых рыб. Для нее характерно отсутствие коллатеральных ганглиев, расчлененность симпатической цепочки и несвязанность с ней черепных нервов. В ряде черепных и туловищных нервов симпатической системы переключение происходит в самом иннервируемом органе, как в парасимпатической системе. Большая часть симпатических нервов имеет короткие преганглионарные волокна, которые оканчиваются в симпатических узлах, лежащих вдоль спинного мозга, и длинные постганглионарные волокна, достигающие иннервируемых органов. У костистых рыб симпатический нервный ствол непрерывен и связан с черепными нервами. Для большинства костистых рыб характерно появление коллатерального ганглия. В их голову начинают проникать туловищные волокна, которые иннервируют ротоглоточную полость и желудок. Из туловищного отдела спинного мозга выходят также нервы, иннервирующие внутренние органы. При этом они имеют короткие преганглионарные и длинные постганглионарные волокна. В каудальной части туловища часть преганглионарных волокон длинные, а постганглионарных короткие. Последние располагаются в стенке мочевого пузыря, что похоже на парасимпатическую систему млекопитающих. Для амфибий и других тетрапод характерна парная симпатическая цепочка ганглиев, идущая вдоль спинного мозга, и многочисленные коллатеральные ганглии, расположенные между внутренними органами. У рептилий и птиц, кроме паравертебрального симпатического ствола, выделяют непарные ганглии: верхний и нижний брыжеечные и чревный (брюшной). Их часто называют превертебральными. Большая часть симпатических ганглиев удалена от иннервируемых органов. Исключением являются ганглии, расположенные в непосредственной близости от половых органов. К этим узлам сигналы приходят по длинным преганглионарным волокнам, а передаются к половым органам – по коротким, что обычно бывает характерно для парасимпатического отдела нервной системы. У млекопитающих парасимпатические ганглии расположены только в области головы и вблизи органов таза. Во всех остальных случаях парасимпатические нейроны лежат в толще и на поверхности легких, сердца и желудочно-кишечного тракта (интрамуральные ганглии). Парасимпатическими нервами млекопитающих, обслуживающими глазные мышцы и железы, расположенные в области головы, являются глазодвигательный, лицевой и языкоглоточный нервы, выходящие из стволового отдела головного мозга. К органам брюшной и грудной полости парасимпатические нервы идут в составе блуждающего нерва. К органам таза парасимпатические нервы идут из крестцового отдела спинного мозга. Афферентные нервные волокна входят в состав крупных периферических нервов. В спинной мозг они входят через спинальные ганглии, однако большая их часть попадает в ЦНС в составе блуждающего нерва. Симпатические постганглионарные нейроны обладают широкой зоной иннервации. Эффект их действия выражается в торможении активности многих внутренних органов и стимуляции сердечной деятельности. Парасимпатическая система действует более локально и оказывает в основном регулирующее действие на функции органов – мишеней. У многих животных в одном и том же органе превалирует либо симпатическая, либо парасимпатическая система. Для низших позвоночных характерно управление многими органами с помощью только одной из систем.
Анатомические преобразования спинного мозга В ряду хордовых животных.
Спинной мозг вместе с головным составляют центральную нервную систему, и граница между ними имеет относительно условный характер, поскольку отростки многих нейронов головного мозга проникают в спинной и наоборот. Границей между этими двумя отделами нервной системы принято считать место перекреста пирамидных пучков нервных волокон. У большинства млекопитающих этот перекрест хорошо виден с вентральной стороны продолговатого мозга. Строение спинного мозга у различных систематических групп позвоночных сильно варьирует. Разнообразие его внешней формы в первую очередь определяется уровнем функциональной нагрузки на конкретную часть этого органа. У четвероногих животных имеют место выраженные утолщения в шейном и поясничном отделах спинного мозга, куда входят нервы, обеспечивающие иннервацию конечностей. У птиц и некоторых млекопитающих в поясничном отделе спинного мозга имеется увеличенная полость, называемая ромбовидным синусом.
|
|||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-06-26; просмотров: 884; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.111.125 (0.067 с.) |