
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Эпигенез и перформизм. Развитие учения о зародышевых листках.Стр 1 из 8Следующая ⇒
Билет 1 Эпигенез и перформизм. Развитие учения о зародышевых листках. ЭПИГЕНЕЗ (от греч. Epigenesis — надрождение, надобразование), учени об эмбриональном развитии как процессе, осуществляющемся путем последовательного появления новообразований и многообразия, в противовес учению о преформации (см.),—предсуществовании в зародыше изначального многообразия.. Термин «Э.» предложен У. Гарвеем (1651), но в полной мере соответствующие взгляды были выражены К.Ф. Вольфом. Вольф исходил из того, что, если правы преформисты, то все органы зародыша, как только мы их можем увидеть, должны быть полностью сформированы. В своей работе "Теория зарождения" (1759 г.) ученый описывает картины постепенного возникновения различных органов из "неорганизованной массы" у животных и растений. К сожалению, Вольф работал с довольно плохим микроскопом, что породило многие фактические неточности, но это не умаляет значения созданной им теории эпигенеза. Преформизм (от лат. «заранее образую») — учение о наличии в половых клетках материальных структур, предопределяющих развитие зародыша и признаки развивающегося из него организма. Преформизм возник на базе господствовавшего в XVII—XVIII вв. представления о преформации, согласно которому зародыш уже сформирован в половых клетках, и его дальнейшее развитие заключается только в увеличении в размерах. Учёные того времени разделились на анималькулистов и овистов. Первые считали, что зародыш содержится в сперматозоидах, вторые — в яйцеклетках. Преформистами были такие ученые, как Антони Левенгук, Марчелло Мальпиги и др. Зародышевые листки – слои тела зародыша многоклеточных животных, образующиеся в процессе гаструляции и дающие начало разным органам и тканям. У большинства организмов образуется три зародышевых листка: наружный — эктодерма, внутренний — энтодерма и средний - мезодерма. Производные эктодермы выполняют в основном покровную и чувствительную функции, производные энтодермы — функции питания и дыхания, а производные мезодермы — связи между частями зародыша, двигательную, опорную и трофическую функции. Учение о зародышевых листках — одно из основных обобщений в эмбриологии — сыграло большую роль в истории биологии. Образование зародышевых листков — первый признак дифференцировки зародыша.
Первым, кто обратил внимание на возникновение органов из зародышевых листков, или пластов, был К. Ф. Вольф. Изучая развитие цыпленка, он показал, что из «неорганизованной, бесструктурной» массы яйца возникают зародышевые листки, дающие затем начало отдельным органам. К. Ф. Вольф различал нервный и кишечный листки, из которых развиваются соответствующие органы. Впоследствии X. Пандер, последователь К. Ф- Вольфа, также описал наличие у куриного эмбриона зародышевых листков. К. М. Бэр обнаружил наличие зародышевых листков и у других животных, в связи с чем распространил понятие о зародышевых листках на всех позвоночных. Так, К. М. Бэр различал первичные зародышевые листки, называя их анимальным и вегетативным, из которых впоследствии, в процессе эмбрионального развития, возникают вторичные зародышевые листки, дающие начало определенным органам. Бесполое размножение у червей. Архитомия и паратомия. Выделяют два варианта бесполого размножения: архитомию и паратомию. В случае архитомии тело червя сперва разделяется на фрагменты, а после достраивает передний и задний концы тела. Паратомия же подразумевает обратную последовательность событий: в ходе этого процесса образуется цепочка сцепленных друг с другом разноимёнными концами тела червей. Билет 2. Происхождение первичных половых клеток. Первичные гоноциты. Представление о зародышевом пути. Первичные половые клетки возникают на ранних этапах эмбриогенеза из энтодермальных клеток вегетативного полюса, как, например, у бесхвостых амфибий, или из энтодермальных клеток желточного мешка, как у всех амниот —пресмыкающихся, птиц и млекопитающих. Первичные половые клетки очень рано отличимы от других клеток по крупным размерам и прозрачной цитоплазме. Половые железы в этот момент только начинают формироваться.
Попав в гонады, первичные половые клетки начинают пролиферировать. Они делятся митозом и называются овогониями. У большинства низших позвоночных овогонии сохраняют способность к делению на протяжении всего репродуктивного периода, так как, например, рыбы за один нерест выделяют тысячи яиц, земноводные — сотни. У высших позвоночных количество яиц, которые вызревают одномоментно, редко достигает 15, обычно их бывает меньше, иногда одно, чем можно объяснить и особенности овогенеза. У человеческих эмбрионов женского пола овогонии размножаются наиболее интенсивно между 2-м и 5-м месяцами внутриутробного периода развития, когда их число достигает примерно 7 млн. К 7-му месяцу многие овогонии погибают, а сохранившиеся входят в профазу первого деления мейоза. Для части яйцеклеток мейоз завершается в момент наступления полового созревания, а для части — непосредственно перед менопаузой. Гоноци́т или первичная половая клетка — эмбриональная клетка, из которой впоследствии могут образоваться сперматозоиды или яйцеклетки. Также гоноцитом могут называться любые клетки, участвующие в процессе гаметогенеза, и сами гаметы. Зародышевый путь - зачатковый путь, ряд поколений клеток от первичных половых клеток (ППК) зародыша до половых продуктов взрослого организма. Вейсман полагал, что в результате неравного в наследственном отношении деления ядра на самых ранних этапах развития особи дифференцируются половые клетки, содержащие, в отличие от клеток тела — соматических, все наследственные потенции яйца; половые клетки образуют непрерывную линию зародышевой плазмы — потенциально «бессмертную» часть организма. Половые клетки обладают рядом морфологических и биохимических особенностей по сравнению с соматическими клетками: у млекопитающих они отличаются высокой активностью щелочной фосфатазы, у некоторых пресмыкающихся — большим количеством полисахаридов и т.д. Билет 3. Регенерация и виды ее. Регенера́ция — способность живых организмов со временем восстанавливать повреждённые ткани, а иногда и целые потерянные органы. Регенерацией также называется восстановление целого организма из его искусственно отделённого фрагмента (например, восстановление гидры из небольшого фрагмента тела или диссоциированных клеток). У протистов регенерация может проявляться в восстановлении утраченных органоидов или частей клетки. Регенерацией называется восстановление организмом утраченных частей на той или иной стадии жизненного цикла. Регенерация, происходящая в случае повреждения или утраты какого-нибудь органа или части организма, называется репаративной. Регенерацию в процессе нормальной жизнедеятельности организма, обычно не связанную с повреждениями или утратой, называют физиологической. Физиологическая регенерация. В каждом организме на протяжении всей его жизни постоянно идут процессы восстановления и обновления. У человека, например, постоянно обновляется наружный слой кожи. Птицы периодически сбрасывают перья и отращивают новые, а млекопитающие сменяют шерстный покров. У листопадных деревьев листья ежегодно опадают и заменяются свежими. Такие процессы носят название физиологической регенерации.
Репаративная регенерация. Репаративной называют регенерацию, происходящую после повреждения или утраты какой-либо части тела. Выделяют типичную и атипичную репаративную регенерацию. При типичной регенерации утраченная часть замещается путём развития точно такой же части. Причиной утраты может быть внешнее воздействие (например, ампутация), или же животное намеренно отрывает часть своего тела (автотомия), как ящерица, обламывающая часть своего хвоста, спасаясь от врага. При атипичной регенерации утраченная часть замещается структурой, отличающейся от первоначальной количественно или качественно. У регенерировавшей конечности головастика число пальцев может оказаться меньше исходного, а у креветки вместо ампутированного глаза может вырасти антенна. Морская звезда регенерирует потерянные лучи. ------------------------------------------------------------------------------------------- Билет 4 Билет 5 Билет №6 Гормоны и рост Гормоны — биологически активные вещества, синтезируемые организмом. Обладая рядом функций и особенностей, гормоны: · управляют жизнедеятельностью организма в целом и являются обязательным компонентом любой его системы; · контролируют генетический аппарат, обеспечивают рост тканей, помогают организму быстро адаптироваться к меняющимся условиям внешней среды; · играют определяющую роль в размножении и развитии потомства; Когда гормон, находящийся в крови, достигает клетки-мишени, он вступает во взаимодействие со специфическими рецепторами; рецепторы «считывают послание» организма, и в клетке начинают происходить определенные перемены. Каждому конкретному гормону соответствуют исключительно «свои» рецепторы, находящиеся в конкретных органах и тканях — только при взаимодействии гормона с ними образуется гормон-рецепторный комплекс. Механизмы действия гормонов могут быть разными. Одну из групп составляют гормоны, которые соединяются с рецепторами, находящимися внутри клеток — как правило, в цитоплазме (стероидные, гормоны щитовидной железы). Будучи жирорастворимыми, эти гормоны легко проникают через клеточную мембрану и начинают взаимодействовать с рецепторами в цитоплазме или ядре. Они слабо растворимы в воде, при транспорте по крови связываются с белками-носителями.
В этой группе гормонов гормон-рецепторный комплекс выполняет роль внутриклеточного реле — образовавшись в клетке, он начинает взаимодействовать с хроматином, который находится в клеточных ядрах и состоит из ДНК и белка, и тем самым ускоряет или замедляет работу тех или иных генов. Биологический результат действия каждого гормона весьма специфичен. В клетке-мишени гормоны изменяют обычно менее 1 % белков и РНК, этого оказывается достаточно для получения соответствующего физиологического эффекта. Большинство других гормонов характеризуются тремя особенностями: · они растворяются в воде; · не связываются с белками-носителями; · начинают гормональный процесс, как только соединяются с рецептором, который может находиться в ядре клетки, ее цитоплазме или располагаться на поверхности плазматической мембраны. В механизме действия гормон-рецепторного комплекса таких гормонов обязательно участвуют посредники, которые индуцируют ответ клетки. Наиболее важные из таких посредников — цАМФ (циклический аденозинмонофосфат), инозитолтрифосфат, ионы кальция. Есть гормоны, у которых внутриклеточный посредник до сих пор не обнаружен (инсулин). Многие исследователи считают, что в таком случае посредниками могут выступать химические соединения, структура которых полностью отличается от структуры уже известных науке посредников. Выполнив свою задачу, гормоны либо расщепляются в клетках-мишенях или в крови, либо транспортируются в печень, где расщепляются, либо удаляются из организма в основном с мочой (адреналин). По химическому строению известные гормоны позвоночных делят на основные классы: · Стероиды · Производные полиеновых (полиненасыщенных) жирных кислот · Производные аминокислот · Белково-пептидные соединения Механизмы действия гормонов различной химической природы имеют сходные черты, так например, гормональные реакции разделяют на начальные, ранние и поздние. Завершением действия любого гормона являются активизация процессов внутриклеточного обмена кальция, сокращения, секреции, энергетического обмена. Гормоны организуют ритмы физиологических функций, в цикле сон-бодрствование, в процессах роста. Кроме этого гормоны синхронизируют суточные ритмы метаболических процессов и осуществляют настройку гормонально зависимых физиологических процессов к факторам внешней среды. Гормон роста (соматотропный гормон, СТГ, соматотропин, соматропин) — один из гормонов передней доли гипофиза. Относится к семейству полипептидных гормонов. Соматотропин вызывает выраженное ускорение линейного (в длину) роста, в основном за счет роста длинных трубчатых костей конечностей. Соматотропин оказывает мощное анаболическое и анти-катаболическое действие, усиливает синтез белка и тормозит его распад, а также способствует снижению отложения подкожного жира, усилению сгорания жира и увеличению соотношения мышечной массы к жировой. Кроме того, соматотропин принимает участие в регуляции углеводного обмена — он вызывает выраженное повышение уровня глюкозы в крови и является одним из контринсулярных гормонов, антагонистов инсулина по действию на углеводный обмен. Описано также его действие на островковые клетки поджелудочной железы, иммуностимулирующий эффект, усиление поглощения кальция костной тканью и др. Значительная часть эффектов гормона роста опосредуется инсулиноподобными факторами роста, главным образом IGF-1 (ранее его называли соматомедином С), который вырабатывается под действием гормона роста в печени и стимулирует рост большинства внутренних органов.
Билет №7 Явление капацитации Капацитация — комплекс последовательных молекулярных изменений на мембране и в цитоплазме сперматозоида, необходимых для индукции гиперактивации и акросомальной реакции. Она завершается в ампуле маточной трубы, которую достигают только несколько сотен (или десятков) сперматозоидов эякулята. Капацитированный сперматозоид способен успешно внедриться в кумулюс и затем связаться с блестящей оболочкой. Кумулюс состоит из фолликулярных клеток и вязкого межклеточного матрикса, в образовании которого участвуют кислые мукополисахариды, гиалуроновая кислота и белки. Эксперименты с клетками животных показали, что кумулюс быстро преодолевают сперматозоиды, находящиеся в состоянии гиперактивации. Внешне она проявляется увеличением интенсивности и амплитуды биений хвоста, что делает движение головки более резким, но менее прогрессивно-поступательным. Такое изменение характера движения сперматозоида не только облегчает ему миграцию через плотную среду, но и увеличивает вероятность его встречи с ооцитом. Время капацитации различно у разных сперматозоидов, что является важной приспособительной реакцией для процесса оплодотворения. Капацитированные сперматозоиды очень активны, однако продолжительность их жизни меньше, чем некапацитированных. Капацитированные сперматозоиды обладают повышенной способностью пенетрировать ткани, что имеет решающее значение в процессе оплодотворения яйцеклетки. У некоторых животных для достижения капацитации необходимо добавление в среду фолликулярной жидкости или клеток яйценосного бугорка. В жидкости спермы кролика, быка, жеребца и других животных, а также человека обнаружен фактор декапацитации, обратимо подавляющий оплодотворяющую способность капацитированных сперматозоидов. Предполагают, что сущность физиологических изменений при капацитации заключается в удалении с поверхности сперматозоидов веществ, блокирующих осуществление акросомной реакции. Билет № 8 Формирование бластены Не похожи на те кл-ки, кот были в этом участке- БЛАСТЕМА. БЛАСТЕМА в учении о регенерации — скопление однородных неспециализир. клеток на раневой поверхности после ампутации органа у животных. В ходе регенерапии из Б. образуются ткани восстанавливающегося органа. В эмбриологии: 1) группа мезенхимных клеток, из к-рых развивается новая особь при бесполом размножении кишечнополостных, червей, асцидий и нек-рых др. животных; 2) поверхностный слой цитоплазмы в центролецитальных яйцах. Билет №9 Моно-, полиспермия. Виды. Сущность оплодотворения состоит в слиянии двух ядер — ядра яйцеклетки и ядра спермия (кариогамия). При слиянии ядра яйцеклетки с ядрами двух и большего числа спермиев происходило бы нагромождение ядерного материала и свойства отцовского и материнского организмов не могли бы наследоваться в равной степени. Поэтому у подавляющего большинства растений и животных оплодотворение идет при участии одного спермия (сперматозоида). Моноспермия - оплодотворение яйцеклетки одной мужской гаметой. Моноспермия - самый обычный способ оплодотворения, вследствие которого яйцеклетка оказывается невосприимчивой к другим гаметам. В очень редких случаях у цветковых растений яйцеклетка может оплодотворяться двумя спермиями (диспермия). Полиспермия 1) у животных — проникновение в яйцо при оплодотворении нескольких спермиев. Различают физиологическую и патологическую полиспермию. Физиологическая свойственна нескольким группам животных с внутренним осеменением (пауки, насекомые, акуловые рыбы, хвостатые земноводные, пресмыкающиеся и птицы). Число спермиев, проникающих в яйца этих животных, варьирует от 1—2 до 10—12 (у насекомых) и нескольких десятков (у хордовых). В ооплазме все спермии изменяются сходно, их головки превращаются в сперматические ядра. Одно из ядер, оказавшееся ближе всего к женскому пронуклеусу, сливается с ним и образует синкарион, который переходит к делениям дробления). Остальные сперматические ядра также вступают в митоз, но у насекомых, хвостатых земноводных и пресмыкающихся он блокируется, и митотические фигуры вскоре резорбируются. У птиц сперматические ядра делятся повторно и дегенерируют на стадиях 8—16 бластомеров; у акуловых рыб они делятся многократно, постепенно вытесняются ядрами дробления (потомками синкариона) за пределы зародышевого диска и не принимают участия в формировании тела зародыша. Подавление митотической активности и резорбция сперматических ядер (кроме одного) обусловлены изменениями свойств цитоплазмы оплодотворённого яйца, природа которых пока неизвестна. Патологическая наблюдается у физиологически моноспермных животных (с наружным и у ряда групп с внутренним осеменением, в том числе у млекопитающих). При слишком высокой концентрации спермиев или плохом физиологическом состоянии яиц механизмы, обеспечивающие в норме моноспермность оплодотворения, недостаточно эффективны, и в яйца может проникнуть по несколько спермиев, которые включаются в развитие, вызывая глубокие нарушения; зародыш рано погибает. Роль полиспермии не совсем ясна, но установлено, что ядро лишь одного из сперматозоидов сливается с женским пронуклеусом. Следовательно, в передаче наследственной информации принимает участие только этот сперматозоид. Ядра других сперматозоидов подвергаются разрушению. Типы дробления 1)По размеру бластомеров. Равномерное и неравномерное. По степени полноты деления Голобластическое дробление. Плоскости дробления разделяют яйцо полностью. Выделяют полное равномерное дробление, при котором бластомеры не различаются по размерам (такой тип дробления характерен для гомолецитальных и алецитальных яиц), и полное неравномерное дробление, при котором бластомеры могут существенно различаться по размерам. Такой тип дробления характерен для умеренно телолецитальных яиц. Меробластическое дробление: Дискоидальное · ограничено относительно небольшим участком у анимального полюса, · плоскости дробления не проходят через всё яйцо и не захватывают желток. Такой тип дробления типичен для телолецитальных яиц, богатых желтком (птицы, рептилии). Такое дробление называют также дискоидальным, так как в результате дробления на анимальном полюсе образуется небольшой диск клеток (бластодиск). Поверхностное · ядро зиготы делится в центральном островке цитоплазмы, · получающиеся ядра перемещаются на поверхность яйца, образуя поверхностный слой ядер (синцитиальную бластодерму) вокруг лежащего в центре желтка. Затем ядра разделяются мембранами, и бластодерма становится клеточной. Такой тип дробления наблюдается у членистоногих. Типы бластул Морула – стадия раннего эмбрионального развития зародыша, которая начинается с завершением дробления зиготы. Клетки морулы делятся гомобластически. После нескольких делений клетки зародыша формируют шаровидную структуру, образованную тесно сближенными бластомерами и не имеющую полости. Например, у гидроидных полипов. Целобластула – стадия зародышевого развития, характерная для некоторых кишечнополостных, низших членистоногих, иглокожих, оболочников, бесчерепных (ланцетник), круглоротых, осетровых рыб и большинства земноводных. Целобластула образуется в результате полного радиального дробления (равномерного или неравномерного); имеет вид пузырька, заполненного жидкостью, однослойная бластула с хорошо выраженным бластоцелем. Стерробластула – стадия зародышевого развития ряда беспозвоночных (некоторых кишечнополостных, червей и моллюсков. Стерробластула образуется обычно в результате спирального дробления яйца. Имеет вид плотного шара; от других типов бластул - целобластулы и дискобластулы - отличается отсутствием полости. Бластула без четко выраженного бластоцеля. Перибластула – стадия зародышевого развития большинства членистоногих животных, имеющих центролецитальные яйца; пузырёк, стенка которого состоит из одного слоя клеток, а полость заполнена нераздробившимся желтком. Образуется в результате поверхностного дробления яйца. Если бластоцель заполнена желтком, т.е. нет типичной полости дробления и бластоцеля. Амфибластула – стадия бластулы у яиц с полным неравномерным дроблением (амфибластические яйца). Такие яйца, имеющиеся у амфибий и некоторых млекопитающих, отличаются большим количеством желтка, скопляющегося на нижнем вегетативном полюсе. Образующаяся в результате дробления амфибластула характеризуется тем, что ее верхняя, анимальная, стенка состоит из мелких бластомеров, тогда как нижняя, вегетативная, образована крупными клетками, богатыми желтком. Полость бластулы лежит эксцентрически, при чем расположена ближе к анимальному полюсу. Целобластула с резко различающимися по размерам бластомерами анимального и вегетативного. Например, губки. Плакула – тип бластулы, характерный для зародышевого развития известковых губок, некоторых червей, асцидий. Имеет вид двуслойной пластинки, образованной более или менее однородными клетками; между слоями — полость дробления. Билет № 10 Билет 11 Гонадотропные гормоны. Гонадотропные гормоны, или гонадотропины — подкласс тропных гормонов передней доли гипофиза и плаценты, физиологической функцией которых является регуляция работы половых желёз. В настоящее время к гонадотропинам относят два гормона передней доли гипофиза: фолликулостимулирующий гормон (ФСГ) и лютеинизирующий гормон (ЛГ), а также особый гормон плаценты — хорионический гонадотропин. Биологическое действие гонадотропных гормонов. Основное действие гонадотропные гормоны оказывают на яичник опосредовано, через стимуляцию секреции гормонов его, благодаря чему создается гипофизарно-яичниковый цикл с характерным колебанием гормональной продукции. Между гонадотропной функцией гипофиза и деятельностью яичника имеется взаимосвязь, играющая основную роль в регуляции менструального цикла. Небольшое количество гонадотропных гормонов гипофиза оказывает стимулирующее действие на гормонопродукцию яичника, вызывая повышение концентрации в крови стероидных гормонов. С другой стороны, значительное повышение содержания гормонов яичника тормозит секрецию соответствующих гормонов гипофиза. Рост и развитие фолликулов, а также секреция эстрогенов стимулируются ФСГ, хотя для полноценной продукции эстрогенов необходимо присутствие и Л Г. Значительное повышение при овуляции эстрогенов тормозит секрецию ФСГ и стимулирует ЛГ, под действием которого развивается желтое тело, секреторная активность последнего усиливается при секреции ЛТГ. Образующийся при этом прогестерон в свою очередь подавляет секрецию ЛГ, и при уменьшенной секреции ФСГ и ЛГ наступает менструация. Эта цикличность в функции гипофиза и яичников составляет гипофизарно-яичниковый цикл, результатом которого являются овуляция и менструация. Секреция гонадотропных гормонов зависит не только от фазы цикла, но и от возраста. С прекращением функции яичника во время менопаузы гонадотропная активность гипофиза повышается более чем в 5 раз, что связано с отсутствием тормозящего действия стероидных гормонов. При этом преобладает секреция ФСГ. Данные о биологическом действии ЛТГ весьма немногочисленны. Считается, что ЛТГ ускоряет рост и развитие молочных желез, стимулирует лактацию и биосинтетические процессы, включая биосинтез белка в молочной железе. Билет 12 1. Гаструляция. Типы. Способы образования мезодермы. Процесс разделения зародыша на зародышевые листки называется гаструляцией, а сам зародыш на этой стадии - гаструлой. Способы гаструляции разнообразны и отчасти связаны со строением бластулы. Иммиграционный тип гаструляции встречается у кишечнополостных. Этот процесс сводится к вселению в полость бластоцеля отдельных клеток, выклинивающихся из стенки бластулы. Когда процесс происходит по всей поверхности бластулы, говорят о мультиполярной иммиграции. Чаще выселение происходит с одного определенного полюса - униполярная иммиграция. Известна также биполярная иммиграция, когда выселение идет с двух противоположных полюсов. У кишечнополостных, где дробление заканчивается морулой без полости, наблюдается другой тип гаструляции, который получил название деламинация (расслоение). В данном случае происходит выравнивание внутренних стенок клеток наружного слоя. Далее вдоль выровненных поверхностей формируется базальная мембрана, отделяющая внешний слой эктодермы от внетренней массы клеток энтодермы. Таким образом, при деламинации клеточные перемещения практически отсутствуют. В целом процесс гаструляции приобретает в ходе эволюции более организованных характер и осуществляется обычно путем инвагинации. В этих случаях внутрь бластоцеля входят не отдельные клетки, а клеточный пласт, не утративший эпителиальной структуры. Полость вворачивания называется гастроцелем, а ведущее в нее отверстие - бластопором (первичным ртом). Края бластопора называются его губами. Так как при инвагинации механическая целостность стенки бластулы не нарушается, вворачивание дна бластулы должно сопровождаться смещением клеточного материала боковых стенок. Такие движения всегда происходят, и скорость их, как правило, не меньше скорости вворачивания. Движения слоя, который в данный момент еще находится на поверхности гаструлы, называют эпиболией (обрастанием). В соответствии с тем типом движения, в результате которого образовалась гаструла, выделяют несколько типов гаструл: 1. инвагинационная 2. иммиграционнная 3. деляминационная 4. эпиболическая Материал, оставшийся на поверхности зародыша после завершения гаструляции - эктодерма. Погрузившийся внутрь листок лишь у кишечнополостных представляет собой чистую энтодерму. У всех вышестоящих систематических групп погрузившийся внутоб материал содержит кроме энтодермы еще и материал мезодермы. Выделяют следующие типы закладки мезодермы: Телобластический (А), встречается у спирально дробящихся форм. В процессе дробления обособляются две крупные клетки, расположенные в полости бластоцеля в области губ бластопора. Они дают начало всей целомической мезодерме личинки. эти бластомеры называются мезобластами или мезотелобластами. Более мелкие мезодермальные клетки отпочковываются от этих крупных бластомеров. В результате возникает пара мезодермальных полосок, которые затем подразделяются на парные сомиты. Внутри сомитов путем расхождения клеток образуются участки вторичной полости тела - целома. Способ образования полостей путем расхождения клеток называется шизоцелевым, или кавитационным. В данном случае мезодерма никак не связана с энтодермой, образующейся из других бластомеров. Принципиально другой - энтероцельный (Б) - способ закладки мезодермы наблюдается у иглокожих и ланцетника. В данном случае материал будущей мезодермы вворачивается вместе с энтодермой в составе гастрального впячивания, которое называется первичным кишечником или архентероном. Мезодерма выделяется из архентерона путем выпячивания его стенок и отшнуровки возникших выпячиваний. Если мезодерма выделяется из архентерона путем деламинации, то выделяют деламинационный (В) тип закладки мезодермы. иммиграционный (Г) тип выделяют в том случае, когда мезодерма отделяется от архентерона путем иммиграции.
Билет 13 1. Провизорные органы, виды и формирование образований провизорных клеток Провизорные органы (нем. provisorisch — предварительный, временный) — временные органы зародышей иличинок многоклеточных животных, функционирующие только в эмбриональный или личиночный период развития. Могут выполнять функции, специфические для зародыша или личинки, или основные функции организма до формирования аналогичных дефинитивных (окончательных) органов, свойственных для взрослого организма. Примеры провизорных органов: хорион, амнион, желточный мешок, аллантоис и серозная оболочка и другие. Амнион — временный орган, обеспечивающий водную среду для развития зародыша. В эмбриогенезе человека он появляется на второй стадии гаструляции сначала как небольшой пузырек, дном которого является первичная эктодерма (эпибласт) зародыша Амниотическая оболочка образует стенку резервуара, заполненного амниотической жидкостью, в которой находится плод. Основная функция амниотической оболочки — выработка околоплодных вод, обеспечивающих среду для развивающегося организма и предохраняющих его от механического повреждения. Эпителий амниона, обращенный в его полость, не только выделяет околоплодные воды, но и принимает участие в обратном всасывании их. В амниотической жидкости поддерживаются до конца беременности необходимый состав и концентрация солей. Амнион выполняет также защитную функцию, предупреждая попадание в плод вредоносных агентов. Желточный мешок —орган, депонирующий питательные вещества (желток), необходимые для развития зародыша. У человека он образован внезародышевой энтодермой и внезародышевой мезодермой (мезенхимой). Желточный мешок является первым органом, в стенке которого развиваются кровяные островки, формирующие первые клетки крови и первые кровеносные сосуды, обеспечивающие у плода перенос кислорода и питательных веществ. Аллантоис - небольшой отросток в отделе зародыша, врастающий в амниотическую ножку. Он является производным желточного мешка и состоит из внезародышевой энтодермы и висцерального листка мезодермы. У человека аллантоис не достигает значительного развития, но его роль в обеспечении питания и дыхания зародыша все же велика, так как по нему к хориону растут сосуды, располагающиеся в пупочном канатике. Пупочный канатик - представляет собой упругий тяж, соединяющий зародыш (плод) с плацентой. Хорион, или ворсинчатая оболочка, развивается из трофобласта и внезародышевой мезодермы. Трофобласт представлен слоем клеток, образующих первичные ворсинки. Они выделяют протеолитические ферменты, с помощью которых разрушается слизистая оболочка матки и осуществляется имплантация. (плод можно сравнить с паразитом) Дальнейшее развития хориона связано с двумя процессами — разрушением слизистой оболочки матки вследствие протеолитической активности наружного слоя и развитием плаценты. Плацента (детское место) человека относится к типу дискоидальных гемохориальных ворсинчатых плацент. Плацента обеспечивает связь плода с материнским организмом, создает барьер между кровью матери и плода. Функции плаценты: дыхательная; транспорт питательных веществ, воды, электролитов; выделительная; эндокринная; участие в сокращении миометрия. Билет 14
Рисунок 1. Поперечное сечение бластоциста и стенки матки через пять дней после оплодотворения. Бластоцист представляет собой полый, наполненный жидкостью шарик, а эта удивительная внутренняя клеточная масса (зеленым цветом) - развивающийся плод. Шарик состоит из клеток трофобласта, который и образуетплаценту. Эндометрий материнской матки (слой розового цвета кровеносными сосудами и желтокоричневый слой поверхностных клеток) готов принять растущий плод и его развивающуюся плаценту. Рисунок 2. Поперечное сечение бластоциста, который имплантируется в стенку матки примерно через шесть дней после оплодотворения. В это время клетки трофобласта постепенно объединяются вместе для того, чтобы образовать синцитиальный трофобласт, состоящий из одной гигантской клетки с множеством ядер. Рисунок 3. Поперечное сечение бластоциста и эндометрия примерно через 12 дней после оплодотворения. Материнская кровь (помечена красным цветом) втекает в смежные пространства, которые развиваются внутри гигантской клетки - синцитиального трофобласта. Эта клетка покрывает поверхность развивающейся плаценты (синим цветом). Кровь плода и его кровеносные сосуды еще не развились. Плод (эмбрион) теперь состоит из двух слоев.
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-06-22; просмотров: 1509; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.140.186.201 (0.101 с.) |





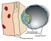 1. Строение бластоцисты (рис)
1. Строение бластоцисты (рис)




