Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
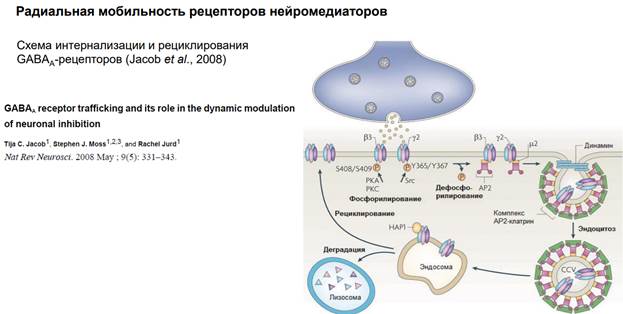
Радиальная мобильность рецепторов нейромедиаторов.
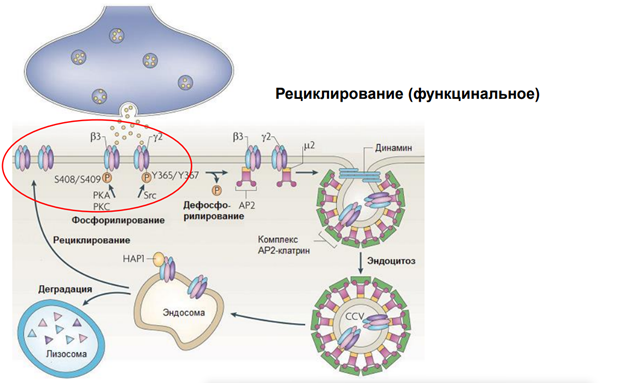
На примере ГАМКа рецептора. Тормозный медиатор. На рисунке изображена пресинаптическая терминаль, выделяется ГАМК на рецепторы. Рецепторы фосфорилированы. Если их дефосфорилировать (оно может быть в результате активации этих рецепторов) (S408/S409 и Y365/Y367 показывают место в субъединице) … АР2 – молекула, выполняющая адаптерную функцию для клатрина. Т.е. рецепторы, когда дефосфорилируются, к ним прицепляется адаптерный белок 2 (АР2) Далее к участку прикладывается молекула клатрина через адаптерный белок и происходит втягивание – эндосомальный процесс … Т.е. несколько рецепторов, которые связались с адаптерным белком, цепляют молекулы клатрина, и утаскиваются внутрь. Эти адаптерные белки сопряжены еще с мембраной (см рис, там они прям на внутренней поверхности мембраны сидят на стадии, когда пошел эндосомальный процесс). В общем, рецепторы утянулсь в просвет полупузырька. Далее белок динамин обеспечивает схлопывание краев пузырька. После этого рецепторы оказываются внутри эндсомы. Такой процесс называется интернализация. Он универсальный. Тут нарисованы ионотропные рецепторы. На самом деле, метаботропные рец-ры также могут удаляться из пресинаптической мембраны.
После этого везикула с рецепторами поглощается эндосомой. Соединяется просто и все. Что с рецепторами? Они могут деградироваться, т.е как бы исчезать. И, собственно, всё.
Тут изображена иная их судьба. У них три судьбы, на самом деле. 1) Деградация, 2) рециклирование… Они могут вернуться в мембрану – рециклирование.
Каким образом еще могут быть получены рецепторы в мембрану? За счет их синтеза. Опускаем тут этот процесс, т.к он требует активации генома, экспрессии генов, которые увеличивают синтез белков, и они встраиваются с помощью эндосом, но берутся они не из мембраны, а с ЭндоПлазматического Ретикулма. Ну и после фосфорилирования новые рецепторы становятся функциональны.
Как можно повлиять на этот процесс? Можно разными способами. Тут опущены транспортные системы, которые обеспечивают такое движение рецепторов.. Поскольку в клетке просто так ничего не плавает. Есть денеины и кинезины, есть актиновые нити.
Здесь остановились в прошлый раз. Циркулирование заключается в процессе интернализации, когда рецепторы захватываются эндосомами. Вот рецепторы, судьба у них разная бывает, на рисунке показана деградация (разрушение), иногда они рециклируют. У деградированного рецептора молекулы уже не функциональны. Если деградация происходит интенсивно, то рецепторы будут исчерпаны и мембрана будет заново составляться. Для этого есть система синтеза, которая обеспечивает этот процесс.
В качестве модели пластичности была вызвана постсинаптическая депрессия.
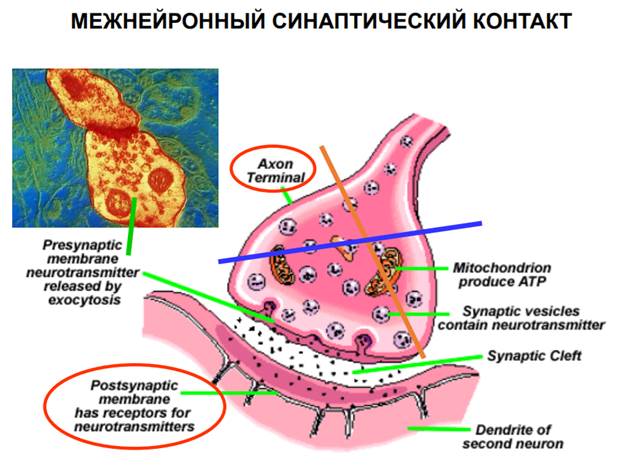
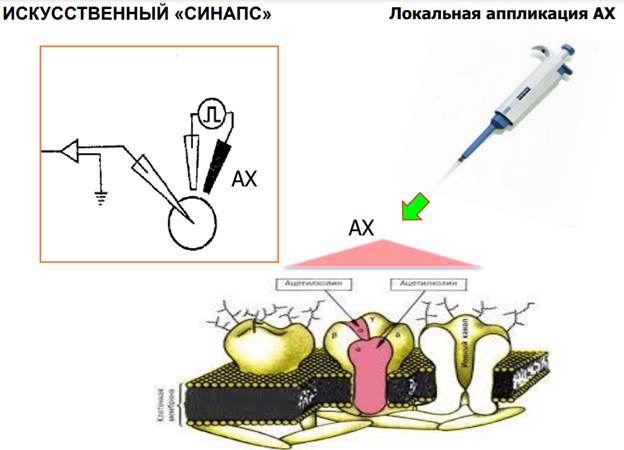
Межнейонный синаптический контакт Искусственный синапс
В этой методике использовался искусственный синапс, для того, чтобы избавиться от мешающих факторов, от которых также зависит пластичность и само постсинаптическое волокно тоже может претерпевать различные изменения и для стандартизации этих условий было решено избавиться от естественного синапса.
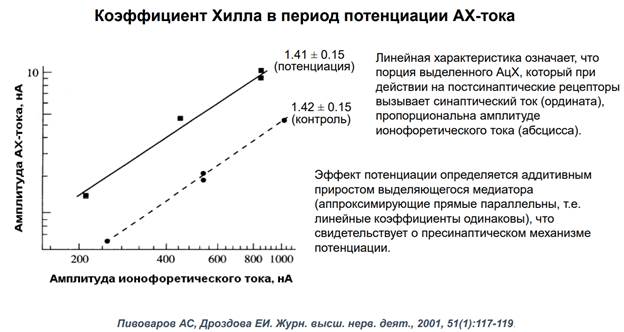
Нужны были контрольные эксперименты. Вот они. Здесь график зависимости А АЦх тока от А ионофоретического тока. Шкала логарефмическая. Ионофорет. ток, котороый выделяется на мембране постсинаптической клетки вызывает активацию АЦх рецепторов. Было проверено на эффекте потенциации, получили увеличении токов. Если ничего не делать (в контроле) получаем такую зависимость как на картинке пунктиром. Количество нейромедиаторов определяется силой тока, который подаем на пипетку. Если слабый ток – выделяется мало АЦх, если много – больше АЦх. Ионофоретический ток дает линейный прирост А тока в нормальных контрольных условиях. А если мы потенциируем нейрон (ч/з пресинапс) и применяем АЦх ток, то мы получаем такие же точки, но выше. Эти апроксимирующие кривые хорошо совпадают, это означает, что мы можем использовать такую технологию для изучения. АЦх действует одинаково, что в норме, что при др воздействиях.
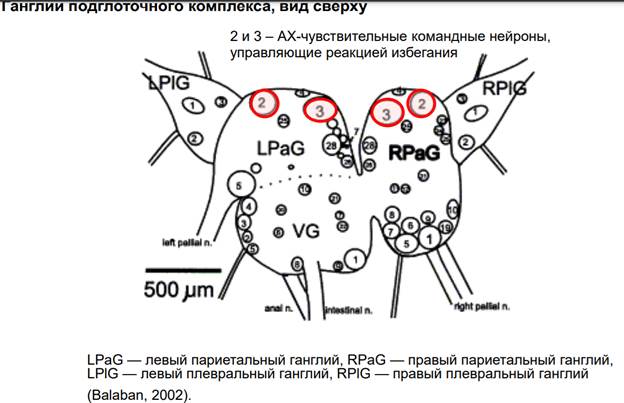
На каких нейронах проводили? На нейронах ганглиев подглоточного комплекса. Эти командные нейроны (которые на картинке) очень хорошо изучены, они управляют реакцией избегании у моллюсков в ответ на воздействия, кот. им не нравятся. Они обладают АЦх чувствительностью (их мембраны содержат АЦх рецепторы).
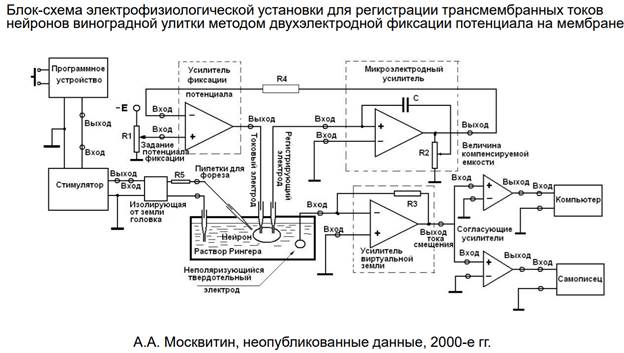
Здесь показана принципиальная схема усилителей, с помощью которых регистрируют трансмембранные токи нейронов виноградной улитки. Здесь двухэлектронная фиксация потенциала на мембране. Мы можем зафиксировать нейрон на любом уровне потенциала ит любые флуктуации мембраны. На таком нейроне можно спокойно регистрировать токи и быть уверенным в чистоте эксп-та. Привыкание
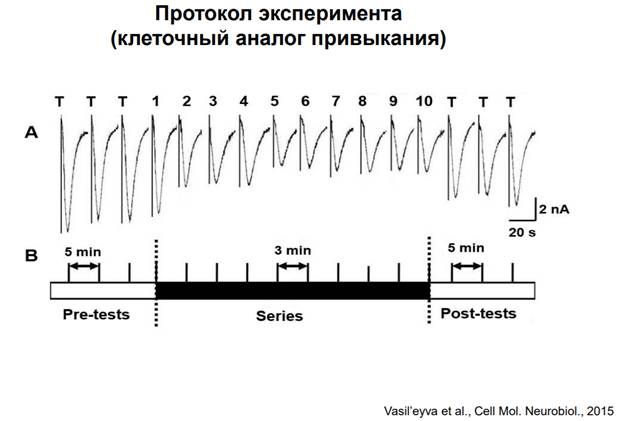
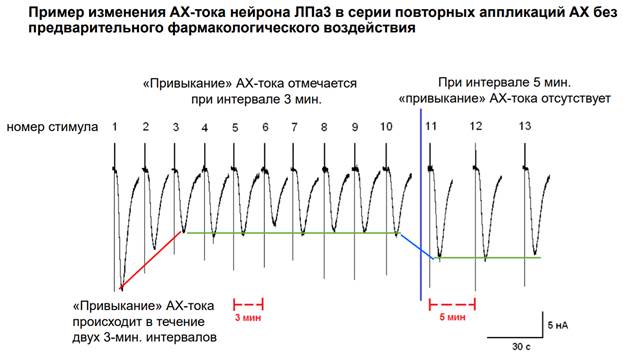
Это одна из работ. Каждая запись - это стимуляция АЦх и запись токов. Вертикальные палки на рисунке – это артефакты. Пики – пиковые значения входящих токов. Нейрон испытывает деполяризацию. Если довольно часто стимулировать синапс, происходит снижении ресурсов для обеспечения токов, может быть и сенситизация рецепторов (говорили об этом выше). Парадигма эксперимента: Дается несколько апликаций АЦх ч/з интервал 5 минут. Здесь есть небольшая депрессия (но статистически она пренебрежимо мала. Затем 10 тестовых стимулов при сниженным интервале 3 минуты. Происходит мощное снижение А токов. Если затем мы снова перейдем на интервал 5 минут, то этого периода хватает, чтобы все токи восстановились. Вот такая модель депрессии.
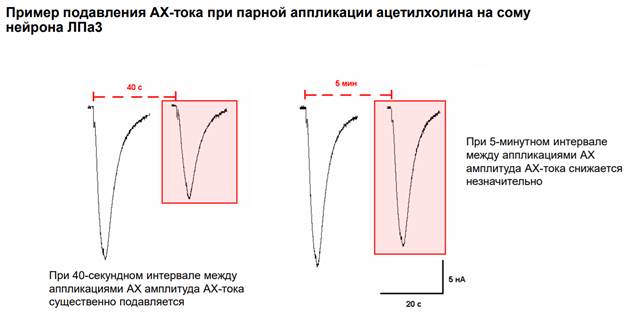
Здесь чуть другие интервалы. Если интервал сделать 40 секунд, то А тока снижается почти в 2 раза. А при интервале 5 минут, снижение незначительное. Т.о. длительные интервалы способствуют восстановлению мембраны, которая реагирует на АЦх. А короткие интервалы вызывают депрессию. Это все медленные процессы.
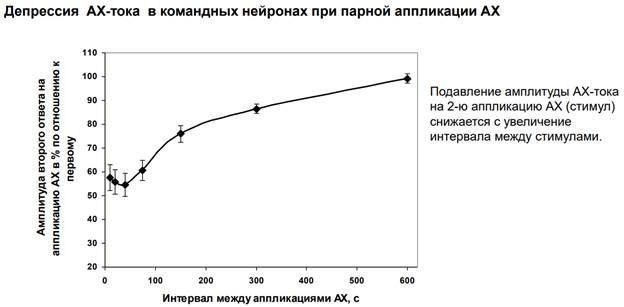
Также здесь были измерены АЦх токи на второй стимул по сравнению с первым стимулом в зависимости от интервалов. За 100% - А на 1ый стимул, остальное отношение чего-то там, он сказал короче говоря нейронный потенциал)). При коротких временах (50 с) мы получаем существенную депрессию. Начинаем увеличивать потенциал – депрессия ослабляется.
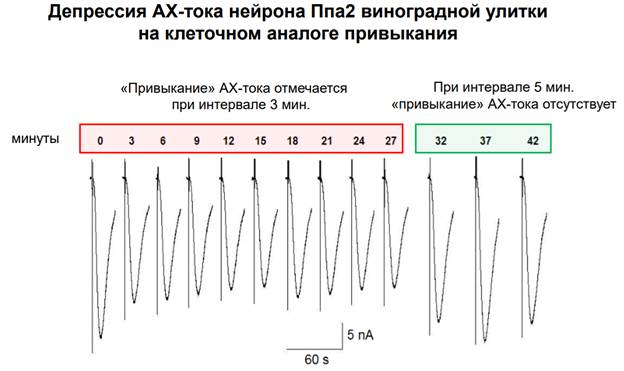
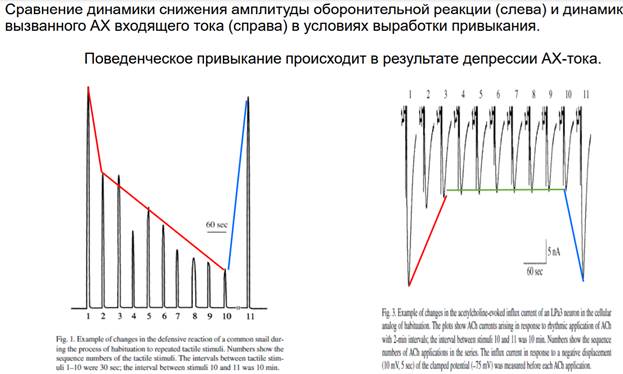
В этой публикации как раз показана депрессия. В ней выделяют 2 фазы: 1) сама депрессия; 2) стабилизация и 3) восстановление. Регистрируется каждый третий стимул. Интервал 3 минуты. В какой-то момент наступает стабильность. Если увеличиваем интервал до 5 минут, А снижается до исходного. Феномен депрессии отражает эффект привыкания (стимул-зависимое обучение=обратный процесс от синситизации). На рис. Выше красное – интервал 3 минуты, зеленое – интервал 5 минут. Поддерживается депрессивное состояние.
Здесь все то же самое, только отмечены интервалы.
Эти измерения показывают, что в основе реакции депрессии снижены токи командных нейронов.
|
|||||||
|
|
Последнее изменение этой страницы: 2022-01-22; просмотров: 78; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.147.238.70 (0.017 с.) |