Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Лекція 6. Енергетична характеристика екосистем
1. Енергетична характеристика середовища. Сонячна енергія як джерело існування наземних екосистем. 2. Концепція продуктивності. Валова, чиста і вторинна продуктивність. Розподіл первинної продукції серед світових екосистем. 3. Потік енергії крізь екосистему. 4. Трофічна структура угруповання. 5. Методи вивчення і виявлення харчових ланцюгів і оцінки швидкості потоку енергії. 6. Екологічні піраміди і енергія харчових ланцюгів. 7. Підтримуюча ємність середовища. 1. Енергетична характеристика середовища. Сонячна енергія як джерело існування наземних екосистем. Кожен організм або екосистема в цілому є відкритою термодинамічною системою, постійно обмінюються з навколишнім середовищем енергією і речовиною відповідно до законів термодинаміки: 1. Енергія переходить з однієї форми в іншу, але не зникає. 2. Ефективність перетворення кінетичної енергії (енергії квантів світла) в потенційну (енергію хімічних зв'язків) завжди менше 100%. Організми, що живуть на земній поверхні, піддаються впливу потоку енергії, що складається із сонячного випромінювання і довгохвильового теплового випромінювання від прилеглих тіл. Сонячне світло, проходячи через атмосферу, послаблюється атмосферними газами і пилом, і в ясні дні поверхні досягає 67% енергії. При цьому ступінь послаблення залежить від довжини хвилі і реальний спектральний склад світла, що досягає поверхні землі в ясний день складається на 10% з ультрафіолетового, на 45% з видимого і на 45% з інфрачервоного світла. Теплове випромінювання від поверхні землі і тіл, температура яких вище 0ºС (ґрунт, вода, живі організми, хмари), на відміну від строго спрямованого вниз випромінювання Сонця, поширюються постійно у всіх напрямках, що може згладжувати коливання енергетичних характеристик середовища за часом доби. Однак, для продуктивності екосистеми і для кругообігу біогенних елементів в ній найважливіше сумарне пряме випромінювання, що потрапляє на ярус автотрофів. Щорічний приплив енергії становить на цей ярус складає 1,1 - 1,5 млрд ккал на м-2 на рік. При цьому 30% енергії відбивається, 46% прямо перетворюється в тепло, 23% витрачається на випаровування і опади, 0,2% витрачається на вітер, хвилі, течії і лише 0,8% витрачається на фотосинтез.
2. Концепція продуктивності. Валова, чиста і вторинна продуктивність. Розподіл первинної продукції серед світових екосистем. Первинною продуктивністю екосистеми називається швидкість, з якою сонячна енергія засвоюється продуцентами і накопичується у вигляді органіки. Виділяють: 1. Валову первинну продуктивність (РG) - загальну швидкість фотосинтезу, включаючи ті органічні речовини, які будуть використані самою рослиною для підтримки свого існування в процесі дихання. 2. Чисту первинну продуктивність (РN) - швидкість накопичення органічної речовини за вирахуванням витрат на дихання. 3. Вторинну продуктивність - швидкість накопичення енергії на рівнях консументів. Вона також ділиться на валову і чисту (мінус трата на дихання). За найсприятливіших умов в валову первинну продуктивність може перетворитися лише 10% загального денного надходження сонячної енергії та за добу до 75-80% валової продукції може перейти в чисту первинну продукцію (в середньому для біосфери ці показники дорівнюють 0,4 і 50% відповідно). Високі швидкості продукування спостерігаються за сприятливих умов і при надходженні додаткової енергії ззовні (за рахунок енергетичних субсидій зменшуються власні витрати на підтримку життєдіяльності). Тому найбільша валова продукція спостерігається в природних угрупованнях, які отримують природну дотацію енергії. Необхідно враховувати, що фактор, який є енергетичної субсидією в одних умовах, може в інших умовах навпаки віднімати енергію у спільноти. Наприклад, посилення транспірації у вологому і в сухому кліматі завдяки вітру. Тобто іноді енергетична субсидія може перейти в стрес. Наприклад, залежність врожайності від концентрації добрив. У межах кожної екосистеми є свій вертикальний градієнт розподілу первинної продукції. Наприклад, в прибережних водах максимум продуктивності на глибині 10 м і зменшується аж до 30 м. Нижче 30 м в прибережних водоймах - слідові кількості продукції. В океанах це 20 і 100 м відповідно. Якщо оцінювати світовий розподіл кількості продукції, то на частку морських екосистем із загальною площею 362 • 106 км² припадає 43,6% загальної валової продуктивності. На частку наземних з площею 135 • 106 км² припадає 57,4%.
Таблиця2. Розподіл екосистем за величиною валової первинної продукції (ВПП).
3. Потік енергії крізь екосистему.
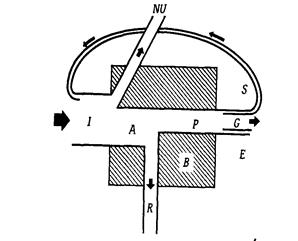
Перенесення енергії їжі від її джерела - автотрофів (рослин) через ряд організмів шляхом поїдання називається харчовий (трофічний) ланцюг. При кожному черговому перенесенні велика частина (80-90%) потенційної енергії втрачається, переходячи в тепло. Організми, які отримують свою енергію від Сонця через однакове число ступенів вважаються такими, що належать до одного трофічного рівня: 1 трофічний рівень - продуценти (зелені рослини); 2 трофічний рівень - первинні консументи (травоїдні); 3 трофічний рівень - вторинні консументи (первинні хижаки); 4 трофічний рівень - третинні консументи (вторинні хижаки). Потік енергії через будь-який компонент екосистеми (окрема рослина, тварина або вся трофічна група) можна представити у вигляді універсальної моделі.
Рисунок 5.Універсальна модель потоку енергії крізь окремий компонент екосистеми (за Одумом):I – енергія, яка надходить на трофічний рівень;NU- невикористана енергія;А – асимільована енергія;Р – продукція;R - дихання;В - біомаса;G – ріст;S - енергія, що накопичилась у запас;Е – екскременти.
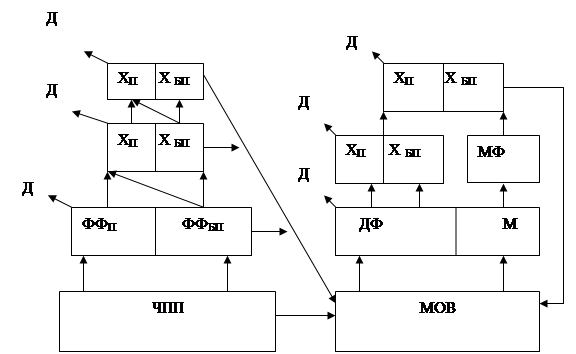
4. Трофічна структура угруповання. У будь-якій наземній екосистемі зустрічаються харчові ланцюги двох типів: пасовищні (починаються з зелених рослин) і детритні (починаючи з мертвої органічної речовини), які разом утворюють двоканальний потік енергії крізь екосистему. Ці шляхи мають ярусне, просторове, часове розмежування. Крім того мікроконсументи (сапрофітні бактерії і гриби) і макроконсументи (фаготрофні тварини) сильно розрізняються відношенням інтенсивності обміну до розмірів організму і вивчаються різними методами.
Рис.6. Узагальнена модель трофічної структури і потоку енергії для наземної екосистеми:ЧПП - чиста первинна продуктивність;МОВ - мертва органічна речовина;ФФ - фітофаги; Х - хижаки; ДФ - детритофаги; М - мікроорганізми; МФ - споживачі мікроорганізмів; Х - хребетні; БП - безхребетні; Д - витрати на дихання.
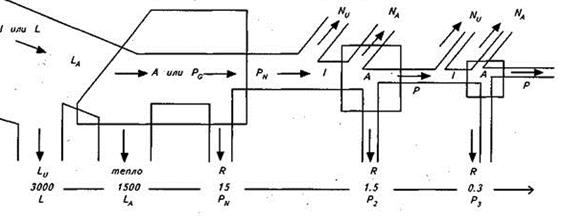
У деяких екосистемах (на пасовищах) по пасовищному ланцюгу може йти 50% і більше потоку енергії. Навпаки, океани і ліси на 90% функціонують як детритні системи. Взаємний зв'язок пасовищного і детритного харчових ланцюгів дозволяє у відповідь на енергетичні впливи перемикати потоки. Однак пряме вилучення травоїдними тваринами і людиною більш 30-50% річного приросту наземної рослинності зменшує здатність екосистеми чинити опір стресу (пасовище стає непридатним за 9 років). Недовипас також може виявитися шкідливим. Якщо пряме споживання живих рослин відсутнє, то детритний матеріал накопичується швидше, ніж йде його розкладання. А це уповільнює кругообіг мінеральних речовин і крім того система може стати пожежонебезпечною. На першому трофічному рівні поглинається близько 50% падаючого світла, а перетворюється в енергію всього 1% поглинутої світлової енергії. Вторинна продуктивність (Р2 і Р3) на кожному наступному трофічному рівні консументів становить в середньому близько 10% попередньої, хоча на рівні хижаків ефективність може бути вище (20%).
В цілому, ефективність переносу багато в чому залежить від поживної цінності джерела енергії (продукти розпаду, подрібнені мікоризними грибами або відмерлі листя і тварини з важко перетравлюваними целюлозою, лігніном, хітином).
Рис.7 Спрощена схема потоку енергії на трьох трофічних рівнях (за Ю.Одумом): I– загальне надходження енергії; LA – світло, що поглинається рослинним покривом; PG – ВПП; A – загальна асиміляція; PN – ЧПП; P – вторинна продукція; NU – невикористана енергія; NA – неасимільована консументами енергія; R – дихання.
Для прогнозування характеру руху енергії по угрупуванню потрібно знати п’ять різних ефективностей переносу: 1. Ефективність поглинання енергії трофічним рівнем (ефективність Ліндемана).
2. Ефективність використання.
Тобто відсоток сумарної продукції одного рівня (Pt-1), який дійсно споживається (з'їдається) наступним трофічних рівнем. Для фітофагів лісів ЕВ приблизно дорівнює 5%, степів - 25%, угруповань з домінуванням фітопланктону - 50%. Для хижаків-хребетних ЕВ 50-100% (якщо їх кормом служать хребетні) і близько 5% - якщо корм - безхребетні. Для хижаків-безхребетних ЕВ дорівнює 25%. 3. Ефективність асиміляції.
Тобто відсоток енергії їжі, що потрапила в травну систему, усмокталась крізь стінки цієї системи і стає доступною для використання. Для мікроорганізмів і грибів, з огляду на позаклітинне травлення, ЕА до 100%, фітофагів і детритофагів - 20-50%, у хижаків - до 50%.
4. Ефективність продукування.
Тобто відсоток асимільованої енергії (At), що включається в нову біомасу (Pt). У безхребетних ЕП висока (30-40%), тому що на дихання витрачається мало енергії. 5. Екологічна ефективність.
Тобто продукція консументів/ продукцію продуцентів
У пойкілотермних тварин – 10%, у гомойотермних – 1-2% (у зв’язку з високою витратою енергії на підтримання температури тіла), у дрібних ссавців нижче (0,86%) завеликих (до 3,14%). При оцінці екологічної ефективності різних трофічних груп степової екосистеми виявилося, що з 100 Дж ПВП 15,2% споживається через систему консументів, а 84,8% - редуцентів, різниця в асиміляції ще істотніше - 9,2% і 90,8% вторинної продукції відповідно. Це результат їх високої активності і здатності переробляти органіку кілька разів поспіль.
Однак консументи в екосистемі, задовольняючи свої потреби в енергії, виконують неоціненну роль регуляторів, зокрема за схемами позитивного зворотного зв'язку на трофічні рівні, що знаходяться вище. Наприклад, а) виїдання рослинності стадами антилоп разом з пожежами під час посухи збільшує швидкість повернення елементів живлення в ґрунт, що в наступний сезон дощів посилює відновлення трави; б) в слині коників накопичується речовина, що стимулює ріст коренів і тим самим збільшує здатність рослини відновлювати листя після поїдання їх коником. 5. Методи вивчення і виявлення харчових ланцюгів і оцінки швидкості потоку енергії. До методів вивчення, виявлення харчових ланцюгів і оцінки обсягу потоку енергії використовують наступні методи: а) спостереження за харчуванням і вивчення вмісту шлунків; б) ізотопні мітки (С13 / С12 - їх різне співвідношення характерно для С3, С4- рослин і водоростей; ці характерні співвідношення передаються і консументам); (Співвідношення Д2 / Н1 в тканинах і фекаліях тварин відповідає цьому співвідношенню в їжі). Використовують радіоактивні мітки і для оцінки швидкості потоку енергії. Наприклад, після додавання в річку Р32 середній час, за який кожна популяція накопичувала максимальну концентрацію радіоактивного фосфору становило: для водних рослин - кілька днів, фільтраторів - 1-2 тижні, для всеїдних - 3-4 тижні, для детритоядних - 4-5 тижнів, для хижаків від 2 до 4 місяців. Розмір врожаю біомаси на корені (загальна суха маса всіх організмів), який може підтримуватися постійним потоком енергії крізь харчовий ланцюг, в значній мірі залежить від розміру особин. Чим менше організм, тим вище його питомий метаболізм і тим менше біомаса, яка може підтримуватися на даному трофічному рівні. Так, врожай бактерій або фітопланктону буде набагато нижче врожаю риби або дерев, хоча ці групи, можливо, використовують однакову кількість енергії, тобто займають однакові трофічні рівні.
6. Екологічні піраміди і енергія харчових ланцюгів. Трофічну структуру екосистеми можна зобразити графічно у вигляді екологічних пірамід, основою яких служить перший рівень (продуценти), а наступні рівні утворюють поверхи і вершину піраміди. Типи екологічних пірамід: а) піраміда чисельності, що відображає чисельність окремих організмів. Форма таких пірамід сильно залежить від розміру продуцентів (фітопланктон або дуби), може бути звернена, що призводить до переоцінки ролі дрібних організмів.
в) піраміда енергії, що показує величину потоку енергії і продуктивність. На форму цієї піраміди не впливають розміри та інтенсивність метаболізму особі, що дозволяє порівнювати екосистему між собою й оцінювати роль популяцій в біотичних спільнотах.
Характер потоку енергії через водну й наземну екосистеми відрізняється за ефективністю і часом, що створює можливість для утворення в водних екосистемах довших харчових ланцюгів. В наземних екосистемах деяка частина енергії швидко розсіюється, що знижує ефективність переносу від одного рівня до іншого, інша ж енергія в протягом тривалого часу зберігається в рослинах та у вигляді детриту. Для розрахунку кількості трофічних рівнів використовують формулу:
Таблиця 3. Розрахунок кількості трофічних рівнів
7. Підтримуюча ємність середовища. Зі збільшення розмірів і складності екосистеми зростає та частка валової продукції, яку спільнота змушене витрачати в процесі дихання на самопідтримку, і падає та частка, яка може йти на подальше збільшення розмірів системи. Частково це компенсується тим, що в суміжних системах виникають додаткові ланцюги і петлі зворотного зв'язку, тобто збільшується ефективність використання енергії та повторного використання речовин і може підвищитися стійкість до порушуючи впливів. Але завжди настає момент, коли витрата і надходження енергії врівноважується, тоді зростання системи припиняється, тобто досягається теоретична максимальна підтримуюча ємність середовища - кількість біомаси, яка може підтримуватися у визначених умовах.
Лекція 7. ЕКОЛОГІЯ ПОПУЛЯЦІЙ 1. Статистичні характеристики популяції. 2. Динамічна характеристика популяції. 3. Структура популяції. 4. Взаємовідносини між популяціями.
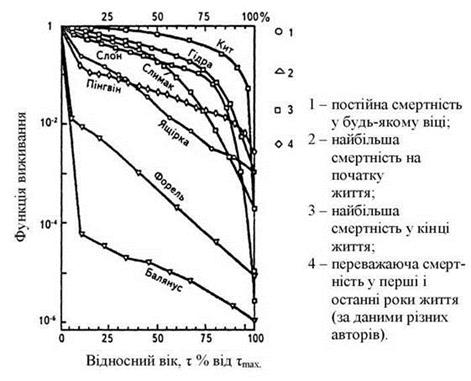
1. Статистичні характеристики популяції. Популяцію можна визначити як будь-яку групу організмів одного виду, що займає певний простір і функціонує як частина біотичного угруповання. Популяція, як і складові її організми, з яких вона складається, диференціюється і підтримує саму себе. Обмін генами перетворює популяцію в відносно цілісну генетичну систему, а взаємини особин дозволяють підтримувати оптимальну в даних умовах чисельність - гомеостаз. Популяціям, як груповим об'єднанням властивий ряд специфічних властивостей: 1) чисельність - загальна кількість особин на всій території; 2) щільність - середнє число особин на одиницю площі (середня щільність - середнє число особин на одиницю всього простору; питома щільність - середнє число особин на одиницю простору, де безпосередньо мешкають особини). Для обліку щільності використовують методи: 1) тотального обліку; 2) пробних майданчиків; 3) методи мічення з повторним виловом; 4) метод без взяття проб (для нерухомих об'єктів); 3) народжуваність - число нових особин, що з'явилися за одиницю часу в результаті розмноження. Розрізняють декілька типів народжуваності: максимальну, яка виявляється в ідеальних умовах, при відсутності факторів, що лімітують; екологічну, що діє при фактичних умовах; абсолютну; питому - на одну особину в популяції (в демографії на 1 жінку репродуктивного віку). 4) смертність - кількість загиблих в популяції особин за певний відрізок часу. Розрізняють екологічну (в даних умовах) і мінімальну (в ідеальних умовах). Часто замість смертності (М) використовують термін - виживаність (1-М). При вивченні популяції складають таблиці виживання, враховують вік особин і питому виживання в кожному віці. На підставі таблиць будують криві виживання.
Форма кривої залежить від турботи про потомство, від щільності популяції. У людей протягом майже всієї історії була висока дитяча смертність, яка різко знизилася з успішним розвитком медицини. Це призвело до зміни типу кривої виживання в поколіннях і стрімкого збільшення чисельності населення Землі, який отримав назву демографічного вибуху.
5) приріст популяції - різниця між народжуваністю і смертністю. Якщо теоретично популяцію не лімітують фактори зовнішнього середовища, то вона здатна до необмеженого росту чисельності і в такому випадку швидкість зростання популяції залежить тільки від величини біотичного потенціалу, властивого виду. Це теоретичний максимум нащадків від однієї особини за одиницю часу.
де
Величина r, наприклад, у козулі = 10-15 потомків, бджоли - 50тис.яєць, риб - до декількох мільярдів ікринок. Різницю між максимальним r і швидкістю зростання, що спостерігається в реальних польових або лабораторних умовах, часто використовують як міру опору середовища, яка характеризує суму всіх факторів середовища, що лімітують і перешкоджають реалізації біотичного потенціалу. 6) темп росту - середній приріст за одиницю часу.
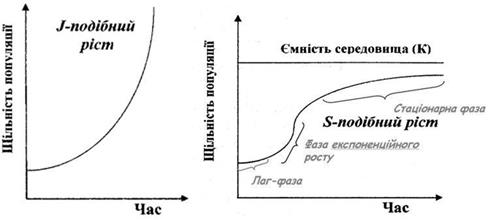
2. Динамічні характеристики популяції. 7) криві росту. Характер збільшення чисельності популяції може бути різним, і в зв'язку з цим виділяють різні типи росту популяції: J - подібний і S - подібний (сігмоїдний). При J - подібній кривій щільність швидко зростає по експоненті, але потім, коли, починають діяти опір середовища або інший фактор, що лімітує, зростання швидко припиняється. Цей тип росту описується рівнянням:
Де При цьому швидкість росту буде постійною, а її величина – пропорційною величині біотичного потенціалу r. J -подібний ріст характерний для видів, ріст чисельності яких не залежить від щільності популяції. Тобто для них відсутні обмежуючі чинники, сила яких проявляється зі збільшенням щільності особин у просторі. Відповідно, в таких популяціях відсутній механізм зворотного зв’язку, який забезпечує саморегуляцію чисельності популяції.
Рисунок 8. Типи росту популяцій.
Такий тип росту чисельності часто є характерним для популяцій при заселенні організмами вільного субстрату (бактерії на поживному середовищі, бур’яни на перелозі), або при виникненні сприятливих кліматичних умов (тепла волога погода, що сприяє розвитку борошнистої роси або фітофтори). Проте жодна природна популяція не може збільшувати свою чисельність нескінченно. У певний момент відбудеться насичення простору особинами, або умови середовища зміняться на несприятливі. Це призведе до обмеження швидкості розмноження, інтенсифікує процеси еміграції особин. Найчастіше у природі при зростанні чисельності організмів включаються специфічні механізми зворотного зв’язку, які починають поступово пригнічувати народжуваність. Такими механізмами можуть бути зменшення трофічних ресурсів, накопичення токсичних метаболітів у середовищі, інформаційні механізми пригнічення розмноження при груповому способі існування. В результаті на певному етапі швидкість росту чисельності популяції знижується. При цьому чисельність популяції стабілізується на певній величині, що відповідає максимально можливій. Ця величина отримала назву ємності середовища – максимальна можлива чисельність популяції, потреби якої можуть бути задоволені ресурсами даної екосистеми. Така крива росту чисельності характеризується S -подібною формою. З математичної точки, така крива описується логістичним рівнянням
Така форма кривої обумовлена поступовим посиленням (з наростанням щільності популяції) дії несприятливих чинників (опір середовища) на відміну від J - образного росту, при якому популяція починає відчувати опір середовища майже в кінці свого росту. Тобто сигмоїдний ріст ще називають ростом, обумовленим щільністю.
8) гомеостаз популяції і регуляція чисельності. Гомеостаз популяції - це підтримка певної чисельності. Регуляція чисельності буває незалежна від чисельності (сильні бурі, раптові падіння температури) або залежна від чисельності (конкуренція, паразитизм і інші біотичні фактори, які призводять у одних видів до загибелі надлишку особин, а у інших до зниження плодючості на основі умовних рефлексів). До жорстких форм можна віднести, наприклад, явище самозрідження у рослин: при великій щільності сходів частина рослин неминуче гине в результаті пригнічення фізіологічно більш сильними сусідами, має значення температура появи сходів, деталі мікрооточення. У рослин регуляція щільності може також здійснюватися шляхом зміни вегетативної потужності кожної особини (відбувається стабілізація не чисельності особин в популяції, а загальної листової фотосинтезуючої поверхні). У тварин жорсткі форми регуляції щільності популяції виявляються лише в тих випадках, коли запаси їжі, води або інших ресурсів різко обмежені (канібалізм у окуня при відсутності інших видів риб; самки наїзників відкладають яйця в дрібних господарів, які представляють собою невеликий запас корму, переважно незапліднені яйця, з яких у перетинчастокрилих розвиваються самці, що впливає на чисельність наступного покоління). Механізмом, що затримує ріст популяції, можуть бути хімічні взаємодії особин (наприклад, один великий пуголовок Rana pipiens може затримати зростання всіх інших в акваріумі на 75 літрів). Інший механізм - прояв при збільшенні щільності інстинктів масової міграції (у попелиць збільшення щільності викликає поява крилатої фази і розліт). Найбільш ефективний механізм стримування зростання чисельності на певній території - територіальна поведінка тварин (мічення і охорона ділянок не допускають розмноження на них чужих особин). Надлишкова частина популяції висиляється, іноді це набуває характеру навалу. При перенаселеності у ссавців в результаті гормональних змін (відповідь на стрес) знижується плодючість, падає народжуваність, підвищується агресивність і рівень смертності. При стабілізації чисельності фізіологічний стан нормалізується.
3. Структура популяції. Популяції властива певна організація: а) просторова структура – характер розподілу особин по території. Розподіл особин в популяції може бути випадковим - 1, рівномірним -2, випадково груповим - 3, рівномірним груповим - 4, груповим, з утворенням скупчення груп. Рівномірний розподіл зустрічається там, де між особинами дуже сильна конкуренція або існує антагонізм, що сприяє рівномірному розподілу в просторі. Розподіл випадкового типу можна очікувати в природі в тих випадках, коли на популяцію одночасно впливають численні, але слабкі фактори. Груповий розподіл виникає в результаті того, що всі особини прагнуть до якогось одного місця.
1 2 3 4 5 Рис.8. Типи просторової структури популяцій.
У кожному конкретному випадку тип розподілу в займаному просторі виявляється пристосувальним, тобто дозволяє оптимально використовувати ресурси. Агрегація може посилювати конкуренцію між особинами за компоненти мінерального живлення, їжу або простір, але це часто більш ніж врівноважується підвищенням життєздатності групи, оскільки група володіє великими можливостями для свого захисту, виявлення ресурсів або зміни мікроклімату. Наприклад, група рослин здатна краще протистояти вітру або ефективніше зменшувати втрату води, ніж окремі особини. Бджоли в вуликах виділяють і зберігають досить тепла для виживання всіх особин при температурі, при якій гинуть окремі особини. Недонаселеність, як і перенаселеність можуть мати обмежуючий вплив. Це принцип Оллі - найбільш сприятлива популяція середніх розмірів. За типом використання простору все рухливі тварини діляться на дві групи: осілі і кочові. При осілому існування тварини протягом більшої частини життя використовує досить обмежену ділянку середовища і часто мають розвинене відчуття дому - хомінг (голуби). Біологічні переваги - на добре знайомій території тварина вільно орієнтується, витрачає менше часу на пошуки корму, найкоротшим шляхом рятується в відомі йому укриття, створює систему запасів (білки створюють серії комор). Однак осілий спосіб життя таїть в собі загрозу швидкого виснаження ресурсів в разі перенаселення. Перевага кочового способу життя полягає в тому, що тварини залежать від запасів корму на конкретній території. Постійні пересування одиночних особин, проте, збільшують ймовірність загибелі від хижаків. Тому кочовий спосіб життя практично не властивий одиночним тваринам, кочують групи, стада, зграї.
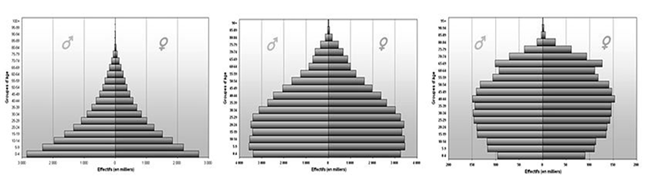
б) вікова структура – співвідношення вікових груп. Для рослин вікова група - це етап онтогенезу, на якому вони характеризуються певними відносинами із середовищем. Так, проростки мають змішане харчування; ювенільні переходять до самостійного харчування, але зберігають просту одновісьову організацію; іматурні рослини мають перехідні ознаки до дорослих вегетативних (типова для виду структура підземних і надземних органів); потім слідує стадія молодих генеративних, середньовікових генеративних, старих генеративних, старих вегетативних (субсенільних), сенільних і відмираючих особин. Співвідношення цих груп називається віковим спектром популяції, який дозволяє виділити наступні типи: - інвазійна (тільки насіння або молоді особини) - нормальна (всі вікові групи), здатна до само підтримки, регресивна (старі генеративні, синільні). У тварин можна виділити три екологічні вікові групи: дореподуктивна,репродуктивна, пострепродуктивна. Співвідношення різних вікових груп (вікова піраміда) в популяції визначає її здатність до розмноження в даний момент і показує, чого можна очікувати в майбутньому: в швидко зростаючих значну частку складають молоді особини; в стабільних віковий розподіл більш рівномірний; в популяції, чисельність якої знижується, буде міститися велика частка старих особин. Розрізняють три типи вікових пірамід: прогресивний, стаціонарний та регресивний. Прогресивний тип характеризується значною часткою в популяції молодих особин і низькою представленістю особин старших поколінь і за формою нагадує трикутник. Такий тип вікової піраміди може характеризувати як позитивні процеси у популяції, коли популяція швидко збільшує свою чисельність, і тому основну частину її складають особини репродуктивного віку та їх нащадки. Проте, подібний тип піраміди буде характеризувати популяції, в яких спостерігається швидке відмирання старших вікових груп. Стаціонарний тип характеризується врівноваженим співвідношенням особин, які складають запас популяції та їх нащадків. Форма піраміди більша нагадує дзвін. Для таких популяцій характерно коли у пари особин в середньому народжується не більше 3-х нащадків. Регресивний тип характеризується переважанням особин старших вікових груп над частою молодих. Форма нагадує кубок. Такий тип не обов’язково відповідає популяціям, що вимирають. Наприклад для організмів, зі високою плодючістю та значною тривалістю життя регресивний тип вікової піраміди є звичайним явищем. Наприклад, у зрілій діброві більшість дерев будуть являти собою старші вікові групи. Молоді дуби будуть зустрічатися як виключення на галявинах, що залишилися після відмирання окремих старих дерев.
Рис. 9. Вікові піраміди
в)статева структура популяції – співвідношення особин різної статі в популяції. Співвідношення чоловічої і жіночої статей в популяції має важливе екологічне значення, оскільки воно безпосередньо пов'язане із потенціалом розмноження, а отже – впливає на життєдіяльність усієї екосистеми. Це пов’язано з тим, що чоловіча особина може запліднити декілька самок. Тому чисельність жіночих особин у популяції має більш важливе значення для існування популяції, ніж її загальна чисельність. У тваринному світі переважають роздільностатеві види, зрідка такі види трапляються і в рослин (тополі, мохи). Також зустрічається явище гермафродитизму, тобто наявність в одному організмі чоловічих і жіночих органів розмноження. Більш характерний для безхребетних тварин та вищих рослин. Існують одностатеві популяції які складаються лише з жіночих особин і розмножуються партеногенезом (попелиці, коловертки, деякі спорові і насіннєві рослини). Як виключення зустрічаються партеногенетичні популяції деяких видів риб, амфібій та рептилій. Співвідношення статей - це відношення кількості самців до кількості самок. Завдяки генетичній детермінації статі у більшості живих організмів кількість самців і самок у хордових тварин майже однакова (1:1). Первинне співвідношення статей визначається сполученням статевих хромосом у процесі мейозу і звичайно буває близько 1:1. Відомо декілька основних шляхів хромосомного визначення статі у тварин. У окремих при сполучень статевих хромосом гетерогаметною статтю виявляються самці, а в інших — самки. Порушення теоретичного співвідношення статей 1:1 при утворенні зигот може бути пов’язано з різницею в інактивації X та Y сперміїв. Статистично доведена менша тривалість існування сперміїв у людини та великої рогатої худоби, які несуть Y-хромосому. Також показано, що у молодих самиць багатьох ссавців частіше народжуються особини чоловічої статі, у середньовікових самиць збільшується частка самок, у самиць старшої вікової групи знову переважають самці. Вторинне співвідношення статей - визначається після народження особин. Варіює у не менш широких межах, ніж первинне. При цьому виявляється, що на вторинне співвідношення статей можуть впливати різні екологічні фактори. Так, наприклад, у багатьох видів черепах, ящірок, крокодилів та змій стать потомства залежить від температури інкубації яєць (температура впливає на активність ферментів, що відповідають за синтез статевих гормонів). Найчастіше низькі температури призводять до народження самців. На визначення статі особини можуть впливати й хімічні фактори. Наприклад, личинка кільчастого черва Bonellia viridis розвивається в самку, якщо після періоду вільного життя вона осідає на дно моря. Якщо ж їй вдається прикріпитися до іншої особини – з неї утвориться самець (який фактично паразитує у статевих протоках самиці, будучи в сотні разів менше за неї). Третинне співвідношення статей – це співвідношення на момент настання статевої зрілості. Звичайно структура популяції може змінюватись за рахунок елімінації тих чи інших статевих груп (наприклад, самці у ссавців живуть менше, ніж самки). У багатьох живих організмів стать визначається не генетичними механізмами, а зовнішнім середовищем. Наприклад, декоративна рослина арізема японська Arisaema japonica – дводомна рослин. Чи буде це жіноча рослин чи буде це чоловіча рослина залежить від маси бульб: найбільші бульби дають рослини з жіночими квітками (бульба містить запас поживних речовин, необхідний для формування плодів).
г) етологічна структура популяції. Закономірності поведінки тварин складають предмет окремої науки – етології. Поведінка тварин по відношенню до інших членів популяції залежить передусім від того, поодинокий або груповий спосіб життя властивий для виду. Поодинокий спосіб життя, за якого особини у популяції незалежні і відокремлені одна від одної, характерний для багатьох видів. Але абсолютно поодинокого існування організмів у природі не зустрічається, тому що при цьому було б неможливим здійснення їх основної життєвої функції – розмноження. Такий спосіб існування вважають властивим для багатьох комах (сонечко, жуки-туруни). Подальше ускладнення відносин всередині популяції призводить до утворення груп: сімей, колоній, зграї і стад. В міру ускладнення таких відносин виникає злагоджена складна поведінкова організація, де кожна особина має свій ранг. Ранг кожної особини у стаді визначається багатьма чинниками. Мають значення вік, фізична сила, досвід і спадкові якості тварини. Більш сильні і досвідчені, зі стійким типом нервової системи, як правило, домінують над більш слабкими. Домінування проявляється в перевазі при споживанні їжі, праві на розмноження, порядку пересування в групі і т. п.
. Лекція 8. ЕКОЛОГІЯ БІОЦЕНОЗІВ 1. Визначення та границі біоценозів. 2. Видова структура біоценозу. 3. Просторова структура біоценозу.
|
|||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-07-19; просмотров: 159; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.191.102.112 (0.126 с.) |

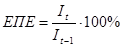 (для першого рівня - це
(для першого рівня - це 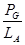 )
)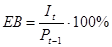
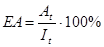
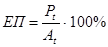
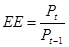
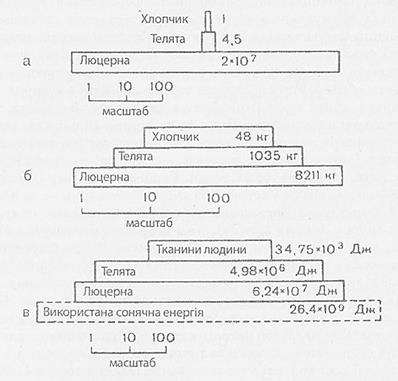 б) піраміда біомаси, що характеризує загальну суху масу, калорійність живого речовини. Так само може бути звернена і призводити до переоцінки ролі великих організмів.
б) піраміда біомаси, що характеризує загальну суху масу, калорійність живого речовини. Так само може бути звернена і призводити до переоцінки ролі великих організмів. 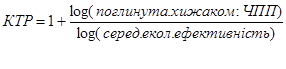
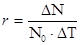
 ,
,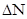 - максимально можливий приріст популяції,
- максимально можливий приріст популяції, 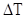 - відрізок часу, N0 – початкова чисельність популяції.
- відрізок часу, N0 – початкова чисельність популяції.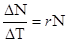 ,
,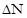 - зміна кількості організмів,
- зміна кількості організмів, 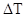 - проміжок часу, N - кількість організмів.
- проміжок часу, N - кількість організмів.