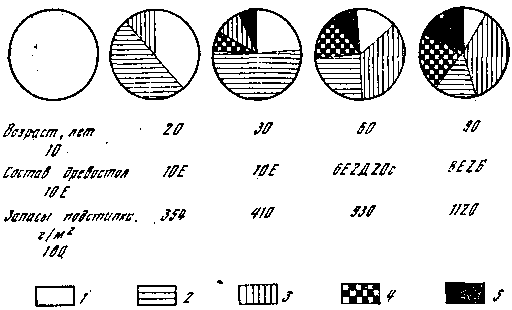Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Другие экологические группы макромицетов
Представители микоризообразователей‑симбиотрофов, напочвенных и дереворазрушающих (ксилотрофы) сапротрофов составляют, как правило, 90–95% видов макромицетов, обитающих в конкретном типе леса, регионе, административном районе грибов. Остальные 5–10% приходятся на пять специализированных экологически и трофически групп: карботрофы, копротрофы, бриотрофы и сфагнотрофы, микотрофы. Формирование этих групп можно рассматривать как результат биохимических адаптаций и ухода от конкуренции со стороны других макромицетов в недоступные для последних условия местообитания. Грибы этих групп изучены крайне слабо, но можно предположить, что их ферментативный аппарат столь же специфичен, как и у грибов других экологических групп. Всеми микологами отмечается четкая (обязательная) приуроченность этих грибов к определенному субстрату и полное их отсутствие на не свойственных для них видах органических веществ. Все названные группы отличаются чрезвычайно бедным видовым составом и небольшой численностью плодовых тел. Карботрофы поселяются на старых кострищах, пожарищах обычно после колонизации их аскомицетами из родов Piziza, Aleuria. После них или одновременно с ними появляются базидиальные макромицеты Pholiota carbonaria, Tephrocybe atrata, T. carbonaria, Fayodia maura. Субстрат, на котором развиваются карботрофы, представляет собой смесь минеральных частиц почвы с обуглившимися остатками древесины, которые являются для грибов богатой питательной средой, поскольку содержат чистый углерод с небольшой примесью полимерных углеводов, скорее всего типа лигнина. Кроме того, можно предположить, что уголь как прекрасный сорбент поглощает в большом количестве продукты жизнедеятельности термофильных грибов и микроорганизмов, поселяющихся в самом начале на этом субстрате, а также обогащается некоторыми минеральными и органическими веществами, поступающими с атмосферными осадками, проходящими сквозь кроны деревьев. Функциональная направленность карботрофов – подготовка специфических местообитаний для последующего заселения их другими организмами, в частности растениями. Копротрофы – это грибы, которые используют в процессе жизнедеятельности органические вещества, находящиеся в экскрементах животных. Для них этот субстрат является единственным источником питания и потому определяет их распространение в природе. Разложение экскрементов травоядных животных на начальных этапах осуществляется низшими грибами, затем их сменяют макромицеты из родов Coprinus, Panaeolus, Stropharia, Conocybe. В процессе эволюции у копротрофных грибов выработался ряд адаптивных признаков, таких, как термофильность спор, стойкость к воздействию ферментов пищеварительной системы животных, способы распространения спор. Ферментативный аппарат грибов этой группы характеризуется широким набором, в котором присутствуют комплексы гидролитических и окислительных ферментов. Экологически субстрат копротрофных грибов имеет особенности, связанные с биохимической спецификой органического материала и с гидротермическим режимом (резкие колебания влажности, постоянно повышенная температура, характерная для компоста). Биохимическая неоднородность и богатство субстрата органическими веществами определяют видовое богатство группировок копротрофов, гораздо более выраженное, чем у карботрофов. Особенно многочисленны грибы на экскрементах травоядных животных, причем набор видов почти не зависит от систематической принадлежности последних. В связи с экологической спецификой субстрата копротрофные макромицеты обладают нежными (хрупкими) плодовыми телами и коротким сроком жизни (несколько дней).
Микотрофные макромицеты относятся к группе сапротрофных микофилов (любящих грибы), развивающихся на мумифицированных плодовых телах шляпочных грибов в большинстве своем из родов «грузди», «сыроежки». Они завершают процесс разложения в природе такого специфического субстрата. Большинство микофилов, насчитывающих около 1500 видов, относятся к микопаразитам и обнаруживают узкую хозяйственную приуроченность, что и обусловливает их экологию и распространение. Ферментативный аппарат микотрофов рассчитан на использование белков и углеводов, составляющих основное содержание плодовых тел шляпочных грибов. Видовой состав микотрофов отличается удивительным однообразием и отсутствием приуроченности к определенным местообитаниям. В лесу довольно часто можно видеть группы маленьких белых грибочков, «уютно» устроившихся на высыхших прошлогодних шляпках черного груздя или свинушки.
Среди бриотрофных макромицетов, разлагающих отмершие части мхов, можно выделить виды, приуроченные к зеленым мхам и сфагновым. Гидрофильность и биохимическая специализация предопределяют их экологию и распространение в природе. К сожалению, ферментативный аппарат грибов этой группы практически не изучен, но их обособленность позволяет предположить определенную специфику в этом плане. Большим видовым разнообразием обладают группировки бриотрофов, приуроченных к сфагновым болотам (виды родов Pholiota, Galerina, Collybia, Mycena). Бриотрофы, как и карботрофы, копротрофы, микотрофы, тесно взаимосвязаны со спецификой субстрата и более или менее независимы от остальных биотических и абиотических факторов среды, т. е. обладают свойствами космополитных организмов. Таким образом, ясно, что макромицеты разделяются на экологические группы, специализированные на разложении биохимически разнородного органического вещества. Изученность групп неравноценна, и кроме того, большая часть исследований проводится в лабороторных условиях. Полученные при этом результаты вряд ли можно с полным правом переносить в природу, где процессы жизнедеятельности грибов проходят в различного рода взаимоотношениях с другими организмами, которые могут либо стимулировать, либо ограничивать эти процессы.
География грибов
Распространение макромицетов на огромной территория Советского Союза исследовано неравнозначно. Очень плохо (на уровне ограниченных районов) изучены грибы севера и центра европейской части СССР, республик Средней Азии, Западной и Восточной Сибири, совсем не изучены Арктика и Крайний Север азиатской части страны. Необходимо отметить и то, что существует прямая зависимость между количеством зарегистрированных в отдельных регионах видов макромицетов и продолжительностью микологических исследований, а также числом микологов, проводящих работы в конкретных районах, областях, республиках. По литературным сведениям, максимальное количество макромицетов – в Приморском крае (802 вида) и в Эстонии (736 видов), наименьшее – на Таймырском полуострове (46 видов), в Туркменской (165) и Казахской ССР (188). Для остальных районов среднее число видов колеблется от 300 до 500. Видовой состав и возраст древостоя, разнообразие типов леса, лесистость и географическое положение района исследований, безусловно, влияют на количество макромицетов и на соотношение экологических групп грибов, однако везде, кроме Крайнего Севера и Юга, на первом месте стоят сапротрофы, а на втором – микоризообразователи, т. е. симбиотрофы. В Таймырских тундрах симбиотрофы составляют 78,5% от всего количества зарегистрированных видов. Учитывая наличие здесь вечной мерзлоты, заболоченности и низкой аэрации болотно‑глеевых почв, крайне неблагоприятных для растений гидротермических условий обитания и широко развитой у кустарниковых форм ив и берез поверхностной корневой системы, можно предположить почти полную микотрофность тундровых растений. Большинство грибов образуют микоризу на корнях карликовой березы, стелящейся по земле, их плодовые тела (березовики, осиновики) по высоте часто превосходят своих симбионтов – деревьев. Короткое северное лето корректирует время появления и продолжительность плодоношения грибов, но масса их достигает внушительных размеров.
При переходе от зоны тундры к лесотундре количество видов макромицетов возрастает довольно резко. Здесь ярко выражена особенность симбиотрофных макромицетов – они теряют облигатность связей с определенной древесной породой. Верные спутники сосны, сопровождающие ее на всей территории распространения, встречаются в лесотундрах Кольского полуострова без нее, т. е. ареалы грибов шире, чем у симбионтов. Набор видов для различных древесных пород, растущих в неблагоприятных климатических условиях на каменистых и щебнистых маломощных почвах, оказывается удивительно однообразным. Так, грибы, растущие южнее, образуют микоризу только с сосной, пихтой, кедром, приурочены к ели. Ряд характерных для дубрав симбиотрофов вступают во взаимоотношения с березой. Таким образом, в крайне неблагоприятных для деревьев условиях обитания обязательность симбиотических связей между партнерами ослабляется вплоть до полного исчезновения и набор видов, образующих микоризу с различными деревьями, становится универсальным. Растения, находясь в экстремальных условиях, теряют жесткость связей с определенными, верными для них, грибами, вступая в симбиоз с теми, которые в силу экологических особенностей могут обитать в данных условиях. При продвижении от лесотундры к тайге и далее к зоне хвойно‑широколиственных лесов количество симбиотрофных макромицетов резко увеличивается, достигая максимума в смешанных лесах. В этом же направлении изменяются почвенно‑грунтовые и гидротермические условия обитания для растений, становясь все более оптимальными, растет видовое разнообразие деревьев и кустарников – потенциальных симбионтов макромицетов. Увеличение количества видов микоризообразователей сопровождается облигатностью связей в отношении симбионта, верностью определенной древесной породе, упорядоченным размещением плодовых тел грибов, уменьшением численности каждого вида и относительно быстрой сменой слоев. Здесь особенно ярко проявляются конкурентные взаимоотношения между видами, принадлежащими к одной экологической группе. Микологических данных для зоны широколиственных лесов недостаточно, чтобы провести объективный анализ распределения симбиотрофных грибов, но по имеющимся сведениям количество их меньшее, чем в хвойно‑широколиственных лесах. Дальнейший спад числа видов микоризообразующих макромицетов наблюдается в лесостепях и степях.
Для искусственных, в частности полезащитных, насаждений степей и полупустынь характерны, как и на Крайнем Севере, отсутствие специфичности видового состава грибов, образующих микоризу с различными деревьями, очень бедный видовой состав и высокая численность грибов, способных плодоносить в крайне неблагоприятных гидротермических условиях названных зон. В горных лесах Молдавии, Крыма, Кавказа и Средней Азии с их высотной поясностью прослеживаются все те закономерности, которые наблюдаются в зональных равнинных лесных сообществах европейской части СССР, т. е. количество видов симбиотрофов увеличивается в зависимости от количества древесных пород и условий обитания для них, а верность грибов определенной древесной породе нарастает в сторону экстремальных пределов произрастания симбионтов. Большую часть территории Сибири и Дальнего Востока занимают леса, в которых процент микоризообразующих грибов несколько снижается по сравнению с лесами европейской части СССР за счет увеличения числа видов, относящихся к другим экологическим группам. Наличие разновозрастных лесных сообществ, находящихся в самых различных по почвенно‑грунтовым условиям местообитаниях, большой набор древесных пород в лесах Дальнего Востока и благоприятная гидротермическая обстановка в них вызывают чрезвычайно выраженное видовое разнообразие симбиотрофных грибов. Сапротрофы включают три основные группы грибов – ксилотрофы, подстилочные и гумусовые макромицеты, составляющие в отдельных регионах до 70% от общего количества видов. На долю остальных групп приходится от 2 до 14 %. Ксилотрофы, т. е. дереворазрушающие грибы, в своем распространении связаны с количеством и качеством субстрата (отмершей древесины). Их видовой состав и обилие являются показателями возраста, лесорастительных свойств и состояния древостоя, интенсивности антропогенных или стихийных (пожары) вмешательств. На фоне меньшего, чем у микоризообразующих грибов, видового разнообразия возрастает число видов с более или менее повсеместным распространением, обладающих широкой экологической пластичностью. В равнинных и горных лесах европейской части СССР, издавна подвергавшихся интенсивной хозяйственной деятельности, ксилотрофы составляют обычно около 20% от общего количества макромицетов, причем на границах распространения лесов видовое разнообразие грибов этой группы резко снижается и сопровождается нарушением обязательной приуроченности к определенному виду субстрата, т. е. сменой либо качества субстрата, либо его состояния. В лесах с благоприятными почвенно‑грунтовыми и гидротермическими условиями (хвойно‑широколиственные, широколиственные) количество дереворазрушающих грибов максимально.
Леса Сибири и Дальнего Востока в сравнении с европейскими характеризуются резким подъемом числа дереворазрушающих грибов вследствие большого распространения старовозрастных древостоев и наличия огромных пространств, подверженных в различное время пожарам, в совокупности приводящих к невероятному количеству отмершей древесины – субстрата для ксилотрофов. На Дальнем Востоке к этим факторам прибавляются благоприятные гидротермические условия приморского климата, а именно высокая относительная влажность воздуха и отсутствие резких перепадов температуры. В связи с этим здесь наблюдается максимальное видовое разнообразие ксилотрофов и самый высокий процент этих грибов в экологической структуре группировок макромицетов. Одна из самых многовидных и специфичных для определенного района группа грибов – подстилочные сапротрофы, которые недостаточно изучены и прежде всего из‑за недолговременности существования плодовых тел грибов и их небольших размеров. Как и для ксилотрофов, ведущим фактором для подстилочных сапротрофов является биохимический состав субстрата – лесной подстилки – и его количество. Но если эти параметры для ксилотрофов ограничены относительно небольшими вариациями (около 300 видов произрастающих на территории СССР деревьев и кустарников, 5–6 градаций состояния субстрата), то для подстилочных сапротрофов число вариаций субстрата во много раз выше. Даже в пределах одного участка леса качественный состав, сложение и запасы подстилки различаются очень существенно. На бесчисленное сочетание вариантов субстрата (соотношение опавшей хвои, листьев, веточек, шишек, отмерших частей травяного, мохового, лишайникового покровов) накладывается чрезвычайное разнообразие микроклиматических особенностей местообитания (температура и влажность субстрата), к которым подстилочные сапротрофы очень чувствительны. Поэтому структура группировок подстилочных сапротрофов в сравнении с другими грунтами грибов наиболее сложная. Поскольку запасы субстрата, зависящего от скорости процессов разложения подстилки, подчиняются зональному распространению, видовое разнообразие подстилочных сапротрофов следует ему. Минимальное количество видов и численность подстилочных сапротрофов наблюдаются в однопородных лиственных лесах, где основная роль в разложении органических веществ, заключенных в подстилке, принадлежит микроорганизмам и почвенным животным. В хвойных лесах, характеризующихся высоким процентом трудноразлагаемых веществ, количество подстилочных сапротрофов увеличивается. Максимума видового разнообразия, а часто и обилия подстилочные сапротрофы достигают в хвойно‑широколиственных лесах. Подстилка здесь мощная, разделена на четкие слои и биохимически разнородная. Учитывая это, в зовах и поясах хвойно‑широколиственных лесов европейской части СССР и Дальнего Востока отмечается самый высокий процент подстилочных сапротрофов. В своем распределении гумусовые сапротрофы подчиняются зональным закономерностям гумусообразования. С севера на юг содержание гумуса в почве, а вместе с этим и количество гумусовых сапротрофов нарастают, достигая максимума в лесостепях, степях, степных поясах гор. Типично лесных видов северных лесов мало, не более 10, с продвижением на юг и появлением в древостое широколи‑ственных пород (дуб, липа, бук, граб) число их возрастает, оставаясь все‑таки значительно меньшим, чем на безлесных территориях. «Лесные» и «степные» гумусовые сапротрофы не имеют общих видов и практически представляют собой разные группы и в видовом отношении, и в отношении условий обитания. Самый большой процент на гумусовые сапротрофы приходится в степях Копет‑Дага, Ленкорани, Белгородской области, т. е. в районах с преобладанием безлесных территорий. Почти такое же количество их на юге Красноярского края, где довольно большие территории заняты остепненными редкостойными лесами и степями. Все остальные группы напочвенных сапротрофов малочисленны и в основном космополитны (повсеместно распространенные) в силу чрезвычайно выраженной приуроченности к субстрату. Это карбо‑, мико‑, гербо‑, копро‑, бриотрофы. Одна из наиболее интересных – группа копротрофов. На них приходится от 1 до 9% от общего количества видов, зарегистрированных в конкретных районах. Видовой состав и численность плодовых тел копротрофов являются индикаторами антропогенного воздействия на растительные сообщества и развития животноводства в регионах, поэтому максимальное их развитие наблюдается в степях Украины, республиках Кавказа и Средней Азии, в некоторых областях Сибири (степных). Таким образом, все группы грибов подчинены в своем географическом распределении определенным закономерностям, связанным в большинстве случаев с зональным и поясным характером распространения растительных сообществ.
Грибы разных лесов
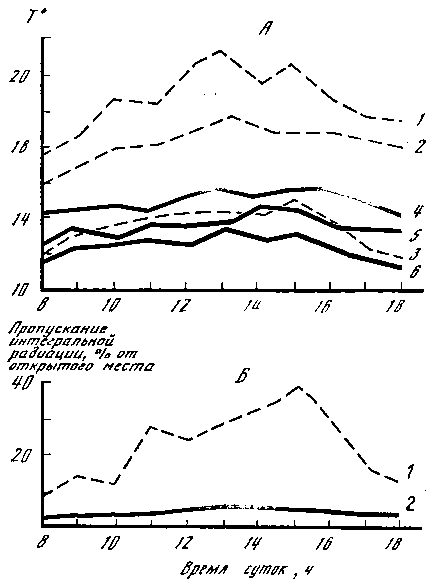
Как формируется видовой состав грибов, соотношение экологических групп, от чего зависит этот процесс? Попробуем ответить на эти вопросы. Молодняки (ельники, сосняки, березняки) в возрасте до 10–15 лет представляют собой такие растительные сообщества, в которых конкуренция за элементы минерального питания, влагу, свет выражена максимально. Практически все деревья здесь находятся в неблагоприятных условиях обитания, за исключением тех, которые растут на опушках. Экологические условия внутри таких массивов относительно однородны. Колебания факторов местообитания по сравнению с лесами более старого возраста незначительны, об этом свидетельствуют данные о суточной динамике освещенности, температуры воздуха и почвы на различных уровнях в 15‑ и 60‑летних ельниках, представленные на рис. 4. В связи с этим относительная влажность воздуха и влажность почвы в молодняках более или менее стабильны в течение суток и вегетационного периода в целом. Соответственно особенностям экологических факторов формируются видовой состав и экологическая структура макромицетов. Последняя чрезвычайно проста и составлена обычно двумя группами – микоризообразователями и подстилочными сапротрофами, приуроченными в своем развитии к слаборазложившейся хвое, листьям, мелким ветвям. Распределение плодовых тел симбиотрофов характеризуется большой скученностью (пятнистостью) и тяготением к границам массивов. Площади пятен не велики (до 10–15 м2), но на каждое плодовое тело в них приходится не более 0,25 м2. Часто наблюдается явление «убегания» плодовых тел симбиотрофов из‑под полога насаждений на расстояние до 5 м и более. Чрезвычайно беден и видовой состав микоризообразующих грибов в молодняках, обычно это один, два вида: в березняках – подберезовик, в ельниках – белый, в сосняках – масленок. Деревья, растущие на границах молодняков, гораздо лучше обеспечены элементами почвенного минерального питания, влагой, светом, чем те, которые находятся внутри массивов. С увеличением возраста последние чаще подвергаются ветровалу, снеголому, что в конечном итоге приводит к их выпадению (гибели) и образованию окон в древесном пологе. Это незамедлительно вызывает перестройку структуры лесных сообществ, вносит разнообразие в условия обитания для грибов. В связи с увеличением запасов подстилки усложняется структура напочвенных сапротрофов, а на выпавших из древостоя породах и больных деревьях появляются ксилотрофы. На место погибших деревьев внедряются другие породы и, естественно, грибы, связанные с ними. Усложнение структуры макромицетов ведет к увеличению их видового разнообразия, но численность представителей всех групп при этом резко падает. Групповое распределение плодовых тел, характерное для молодняков, сменяется рассеянным.
Рис. 4. Варьирование температуры и освещенности в 15‑ и 60‑летних ельниках А – температура в 60‑летнем (1 – на высоте 1,5 м; 2 – на поверхности почвы; 3 – на глубине 5 см) и 15‑летнем ельнике (4 – на высоте 1,5 м; 5 – на поверхности почвы; 6 – на глубине 2–3 см). Б – пропускание интегральной радиации (в % от открытого места): 1 – в 60‑летнем; 2 – в 15‑летнем ельнике
Максимальное количество видов грибов наблюдается в средневозрастных (60–90‑летних) лесах: ельниках, дубравах, березняках, сосняках. Это можно проследить на примере ельников: в 10‑летних общее количество видов макромицетов – 6, в 20‑летних – 16, в 30‑летних – 27, в 60‑летних – 90. Дальнейшее увеличение возраста лесов приводит к уравниванию экологических условий под пологом леса, уменьшению видового разнообразия и численности грибов до определенного уровня, который остается более или менее стабильным продолжительное время. Мы рассмотрели в общих чертах закономерности формирования структуры группировок макромицетов в различных по возрасту и составу лесах умеренного климата. Существенную поправку в эту модель вносят почвенногрунтовые условия, в которых находятся леса. Для древесных пород, занимающих более или менее сходные по обеспеченности элементами минерального питания и влажностью местообитания, эти поправки не велики, в противном случае они достигают огромных величин. Так, например, из найденных в четырех типах сосняков Прикамья 170 видов макромицетов общими для них оказались всего 8, а для такого же числа типов ельников из 180 видов выявлено 83 общих. В формировании видового состава макромицетов играет основную роль не только (и даже не столько) состав древостоя, сколько условия, в которых находятся лесные сообщества. В географически удаленных (Подмосковье и Эстония) ельниках, находящихся в одинаковых почвенно‑грунтовых условиях, видовое сходство грибов составляет 80%. Рассмотрим влияние условий обитания, в которых может произрастать одна и та же лесообразующая порода, на формирование видового состава, численность и структуру микоризообразующих грибов, энергетически непосредственно связанных с деревьями. Как уже отмечалось, деревья в молодняках находятся в крайне неблагоприятных условиях обитания. Низкая освещенность, острая конкуренция за элементы минерального питания и влагу препятствуют процессу микоризообразования, поэтому симбиотрофы в них характеризуются бедным видовым составом, групповым распределением плодовых тел и большой численностью. Эти особенности микоризообразующих грибов характерны не только для молодняков, но и для лесов более старого возраста, находящихся в чрезвычайно неблагоприятных условиях местообитания. К таковым можно отнести березняки, растущие на скалах Кольского полуострова, березовые полезащитные полосы полупустынь Прикаспия, сильновытоптанные березняки Подмосковья. Показатели видового сходства грибов для таких лесов гораздо выше, чем для лесов, образованных той же породой, но находящихся в более благоприятных почвенно‑грунтовых условиях. Переход к более оптимальным условиям произрастания для деревьев сопровождается возрастанием количества видов симбиотрофов, уменьшением обилия плодовых тел каждого вида, отсутствием группового распределения плодовых тел, быстрой сменой во времени грибных слоев.
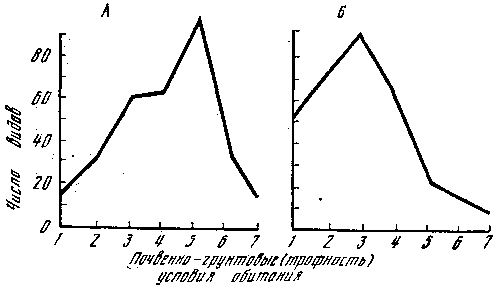
Рис. 5. Распределение симбиотрофов в сосняках и ельниках различного типа А – сосняки; 1 – лишайниковый; 2 – вересковый; 3 – скальный; 4 – бруснично‑зеленомошный; 5 – чернично‑зеленомошный; 6 – кисличный; 7 – долгомошный и сфагновый. Б – ельники: 1 – брусничный; 2 – скальный; 3 – черничный; 4 – кисличный; 5 – черничный влажный; 6 – долгомошный; 7 – хвощово‑сфагновый
Для лесов Карелии (ельники, сосняки) одинаково низкое количество симбиотрофов отмечено как в условиях недостаточной (сосняк лишайниковый, скальный), так и избыточной (сосняк долгомошный и сфагновый, ельник долгомошный, хвощово‑сфагновый) почвенной влагообеспеченности, с которыми связана и обеспеченность растений минеральным питанием (рис. 5). Видовое сходство гораздо выше, например, между ельником и сосняком брусничным (0,74), чем между сосняками лишайниковым и сфагновым (0,37), т. е. в лесах, сложенных одной и той же породой, но находящихся в резко различных условиях обитания. Показательны в этом отношении сосняки, поскольку они занимают самые различные условия обитания и интразональны в своем распространении, но приурочены в основном к бедным песчаным почвам и потому особо нуждаются в симбиотрофных грибах. Обычно с ними связаны 50–70 видов микоризообразующих макромицетов, но в условиях произрастания на богатых меловых почвах их число снижается до 11. В искусственных разновозрастных сосняках, посаженных на богатых суглинистых почвах Подмосковья, встречаются только два вида, верных сосне, – поздний и зернистый масленок, и те в небольшом количестве и не ежегодно, по‑видимому, в таких благоприятных условиях (почвенно‑грунтовых) сосна не нуждается в грибном симбионте. Сосняки на песках обычно образуют светлые редкостойные леса. Хорошая освещенность в сосновых лесах стимулирует отток углеводов к корням деревьев, и интенсивность микоризообразования в данном случае находится в прямой зависимости от концентрации в почве азота, фосфора и калия, их связанности почвенным поглощающим комплексом (доступности для растений) и обеспеченности почв кислородом. Известно, что в ряде почв, особенно при недостатке и избытке влаги, фосфор и калий, непосредственно участвующие в превращении энергии и входящие в состав основных продуктов жизнедеятельности деревьев, прочно связываются почвенным поглощающим комплексом и тем самым исключаются из биологического круговорота. Известно также, что периоду всасывания питательных веществ микоризными корнями предшествует процесс обмена веществ, зависящий в первую очередь от температуры и обеспеченности почв кислородом. Поглощенные корнями деревьев доступные соединения элементов питания поступают в гифы грибов. Фосфор переходит в мицелии в связанное состояние и впоследствии транспортируется в ткани дерева. После отмирания микоризных корней фосфор возвращается в почву. Наибольшее варьирование подвижного фосфора (Р2O5) наблюдается в самых верхних горизонтах почвы – зоне максимальной концентрации тонких корней. Другие доступные для растений элементы минерального питания (в частности, обменные основания), как и фосфор, обнаруживают четкую аналогию с распределением по вертикальному профилю почв гифов микоризообразующих грибов. Например, на сухих почвах под лишайниковым бором гифы достигают глубины 25 см, а в сосняках брусничных, занимающих обычно склоны песчаных холмов (транзитные формы рельефа), а именно верхние их части, где резко выражена скорость транзита минеральных и органических веществ с водой, гифы проникают на глубину 160 см и успевают перехватить большую часть этих элементов для своих партнеров‑деревьев. Наименьшая обеспеченность растений доступными соединениями фосфора отмечена на песчаных почвах повышенных (лишайниковые боры) и пониженных (сфагновые сосняки) форм рельефа. Однако скорость транспорта питательных веществ из мицелия грибов в ткани растений на бедных почвах гораздо выше, чем на богатых.
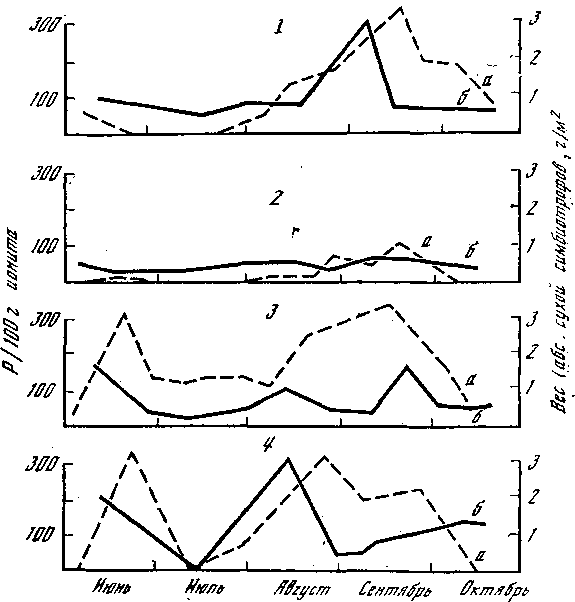
Рис. 6. Динамика фосфора в горизонте А1 дерново‑подзолистых почв в массы карпофоров симбиотрофов вблизи эдификатора 1 – ель; 2 – дуб; 3 – береза; 4 – окно; а – фосфор; б – симбиотрофы
Немаловажным фактором для микоризообразования является и обеспеченность почв кислородом, но грибы приспособились к его недостатку путем образования водонепроницаемой пленки на гифах мицелия в условиях почвенного переувлажнения и дефицита влагообеспеченности местообитаний. Время появления и количество плодовых тел грибов‑микоризообразователей, к которым относятся большинство съедобных и ядовитых (березовики, осиновики, грузди, сыроежки, маслята, белые, лисички, мухоморы), в сильной степени зависят от содержания фосфора в лесной подстилке и верхних почвенных горизонтах, а также от его динамики. Максимальные их количества совпадают во времени и в пространстве, о чем свидетельствуют данные, представленные на рис. 6. Работы проводили в смешанном лесу, где отдельные участки находились под влиянием различных пород (эдификаторов), а были такие, где породы древесные выпали из древостоя и образовались окна в древесном пологе. В таких окнах, которые можно считать аналогом лесных опушек, наблюдается два пика максимума, соответствующих раннелетнему и осеннему слоям грибов. Дуб в условиях Подмосковья – маломикотрофная порода, т. е. с ним связано небольшое количество симбиотрофов, и соответственно этому содержание фосфора (Р) под ним имеет вид выровненной кривой. Способность образовывать микоризу на своих корнях выражена у различных деревьев неодинаково, с одними связаны десятки видов, с другими – единицы. Некоторые виды грибов верны своим партнерам и сопровождают деревья на всем протяжении их географического распространения, что и закреплено в названии грибов (осиновик, березовик, лиственничный масленок, сосновый рыжик); другие не так верны, древесной породе. Совсем нет симбиотрофов у ив, черемухи, рябины, липы, клена, вяза, ясеня. С грабом связаны 2 вида, с ольхой, как и с осиной, – 4, с лиственницей – 8, с пихтой – 27. Иначе обстоит дело с древесными породами, которые в силу своих биологических и экологических особенностей могут занимать крайне различные по почвенно‑грунтовым и гидротермическим условиям местообитания. К ним относятся: береза, ель, дуб, сосна. Их способность образовывать микоризу зависит в первую очередь от обеспеченности почв минеральными элементами, доступности для растений питательных веществ и гидротермического режима региона. Показательны в этом отношении березовые леса, распространенные по всей территории СССР от Крайнего Севера до Юга и с Запада до Приморского края. На Севере, где наблюдается лимит или недоступность для корней азота, фосфора, калия, береза чрезвычайно высокомикотрофна и с ней связано около 100 видов симбиотрофов. По мере улучшения почвенно‑грунтовых условий ее микотрофность резко падает, достигая минимума на богатых почвах умеренного климата (30–40 видов). С дубом связано от 10 до 100 видов симбиотрофных макромицетов в зависимости от условий обитания, а с елью – 30–70. Мы рассмотрели, как ведут себя в различных лесах симбиотрофы, непосредственно связанные с корнями древесных пород, а теперь проследим, как ведут себя напочвенные сапротрофы. Формирование их структур в лесных сообществах подчинено закономерностям накопления и разложения мертвого органического вещества, заключенного в лесном опаде и подстилке. Это можно проследить на рис. 7, где представлена схема соотношений различных групп напочвенных сапротрофов в связи с изменениями качественного состава и количества опада и подстилки в возрастном ряду ельников. В молодняках происходит постепенный рост запасов лесной подстилки и процесс ее вертикальной дифференциации (формирование слоев). К 30 годам в ельниках усложняется ее состав, формируется слой погребенных в подстилке трудноразлагаемых фракций (кора, ветки, шишки) и гумусовый слой. С ними связаны соответствующие виды грибов. При внедрении в ельники лиственных пород (дуб, осина, береза) запасы лесной подстилки возрастают. Кроме того, присутствие в опаде листьев существенно меняет темпы разложения хвойной подстилки и влечет за собой появление грибов, специализированных на разрушении мертвого органического вещества лиственных деревьев. Необходимо учитывать и то, что в более старых по возрасту и сложных по набору древесных пород лесах создается огромное количество экологических ситуаций в отношении микроклимата, что вызывает рост видового разнообразия напочвенных сапротрофов.
Рис. 7. Формирование структуры напочвенных сапротрофов в возрастном ряду ельников 1 – сапротрофы на опаде; 2 – сапротрофы слоя L; 3 – сапротрофы слоя F; 4 – сапротрофы погребенных фракций опада; 5 – гумусовые сапротрофы; Е – ель; Д – дуб; Ос – осина; Б – береза
Дереворазрушающие сапротрофные макромицеты наиболее независимы в отношении фитоклиматических особенностей определенного лесного сообщества. Количество видов и их численность зависят от запасов отмершей древесины и ее состояния разложения. С увеличением возраста древостоя, количества древесных пород, образующих леса, масса мертвого органического вещества, заключенного в древесине, растет, соответственно увеличивается и видовое разнообразие ксилотрофов. В среднем для лесов СССР около 40% всех макромицетов принадлежат к группе микоризообразующих, 20% – ксилотрофов и 40% приходится на напочвенные сапротрофы.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-07-19; просмотров: 85; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.135.197.201 (0.041 с.) |