Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
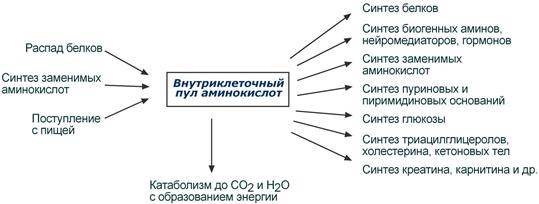
Аминокислотный фонд клетки. Источники пополнения. Пути использования аминокислотного фонда. Заменимые и незаменимые аминокислоты. Механизмы синтеза аминокислот.⇐ ПредыдущаяСтр 15 из 15
Аминокислоты в клетке составляют динамичный пул, который непрерывно пополняется и так же непрерывно расходуется. Существуют три источника аминокислот для пополнения этого пула – поступление из крови, распад собственных внутриклеточных белков и синтез заменимых аминокислот. Путь дальнейшего превращения каждой аминокислоты зависит от вида и функции клетки, условий ее существования и гормональных влияний. Спектр веществ, получаемых клеткой из аминокислот, чрезвычайно широк.
Реакции превращения аминокислот в клетке условно можно разделить на три части, в зависимости от реагирующей группы: - с участием аминогруппы - здесь подразумевается удаление от аминокислоты аминогруппы тем или иным способом, в результате чего остается углеводородный скелет, - по боковой цепи (радикалу) - происходит использование углеродного скелета для синтеза глюкозы, жиров, или для образованеия энергии АТФ, - по карбоксильной группе - связано с отщеплением карбоксильной группы. Заменимые аминокислоты (аланин, аспарагиновая и глутаминовая кислоты и их амиды, аспарагин и глутамин) образуются в результате трансаминирования из промежуточных метаболитов — 2-кетокислот. Пролин синтезируется в достаточных количествах из глутамата, а представители серинового семейства (серин, глицин и цистеин) сами являются естественными метаболитами организма животных. Незаменимые аминокислоты должны поступать с пищей. Так, организм высших организмов не способен синтезировать ароматические аминокислоты de novo (тирозин не является незаменимой аминокислотой только потому, что может образоваться из фенилаланина). К незаменимым аминокислотам принадлежат аминокислоты с разветвленной боковой цепью: валин и изолейцин, а также лейцин, треонин, метионин и лизин. Гистидин и аргинин являются незаменимыми для крыс, но касается ли это также человека — спорно. Трансаминирование аминокислот – перенос аминогруппы (NН2-) от аминокислоты на α-кетокислоту без промежуточного образования аммиака. Реакции трансаминирования катализируют ферменты – аминотрансферазы (или трансаминазы). Кофермент аминотрансфераз – пиридоксальфосфат (производное витамина В6). В реакции принимает участие альдегидная группа кофермента. Реакция легко обратима.
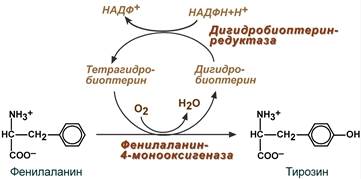
--- 40. Общие пути метаболизма аминокислот. Особенности обмена отдельных аминокислот на примере обмена фенилаланина и тирозина
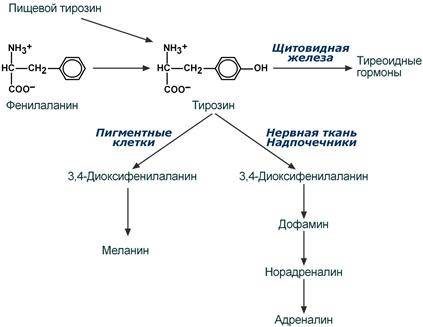
Фенилаланин относится к незаменимым аминокислотам, поскольку ткани животных не обладают способностью синтезировать его бензольное кольцо. В организме фенилаланин используется только в синтезе белков, весь неиспользованный запас аминокислоты превращается в тирозин. В этом непосредственно участвует фермент фенилаланин-4-монооксигеназа (фенилаланингидроксилаза), обеспечивающий окисление ароматического кольца. Кофермент тетрагидробиоптерин в реакции окисляется до дигидроформы. Восстановление кофермента осуществляет дигидробиоптерин-редуктаза с своим коферментом НАДФН.
Тирозин, помимо участия в синтезе белков, является предшественником гормона надпочечников адреналина, медиаторов норадреналина и дофамина, гормонов щитовидной железы тироксина и трийодтиронина и пигмента меланина.
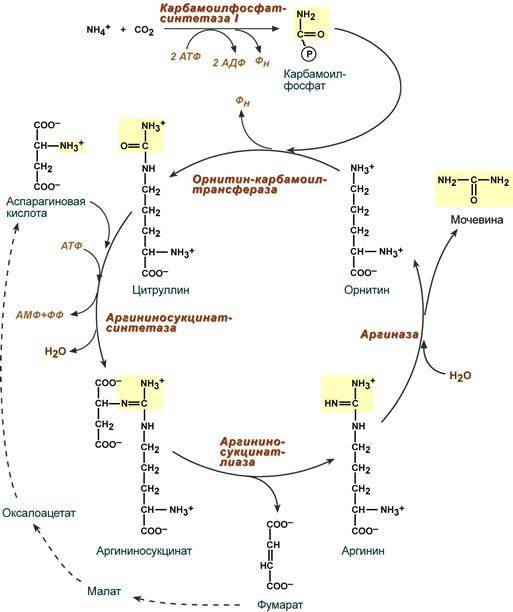
--- 41. Образование мочевины. Роль печени в мочевинообразовании. Другие пути обезвреживания аммиака. Значение исследования уровня мочевины и остаточного азота в клинической практике. В печени весь удаляемый аммиак используется для синтеза мочевины. Увеличение синтеза мочевины наблюдается при распаде тканевых белков и азотистых соединений (голодание, воспалительные процессы, сахарный диабет) или при избыточном белковом питании. У младенцев и детей синтез мочевины может быть снижен по двум причинам: незрелость печени и активный синтез белков и нуклеиновых кислот при росте организма. Определение концентрации мочевины в крови является ценным диагностическим показателем. Реакции синтеза мочевины являются циклическим процессом и получили название орнитиновый цикл. Синтез мочевины начинается в митохондриях (первая и вторая реакции), оставшиеся три реакции идут в цитозоле. Для переноса цитруллина и орнитина через митохондриальную мембрану существуют специальные переносчики. В образовании одной молекулы мочевины участвует 1 молекула NH4+, 1 молекула CO2, аминогруппа 1 молекулы аспарагиновой кислоты, затрачивается 4 макроэргических связи трех молекул АТФ.
Мочевинообразование - одна из важнейших дезинтоксикационных функций печени, заключающихся в преобразовании ядовитых осколков белковой молекулы в нетоксичное вещество. При дезаминировании а/к-т, нуклеотидов и др.азотистых соединений печень принимает участие в синтезе аммиака, превышение предельно допустимых концентраций которого становится высокотоксичным для организма. Дезинтоксикация аммиака происходит посредством использования его для синтеза мочевины. Для удаления аммиака из организма используется включение его в состав мочевины в печени и выведение ее с мочой, и удаление почками в виде аммонийных солей. Однако, так как аммиак является чрезвычайно токсичным соединением, то предварительно в тканях (!) происходят реакции его обезвреживания (временного связывания) для переноса в печень и почки. – синтез глутаминовой кислоты и глутамина, синтез аспарагина: - синтез глутаминовой кислоты (восстановительное аминирование) – взаимодействие α-кетоглутарата с аммиаком. Реакция по сути обратна реакции окислительного дезаминирования, однако в качестве кофермента используется НАДФН. Происходит практически во всех тканях, кроме мышечной, но имеет небольшое значение, т.к. для глутаматдегидрогеназы предпочтительным субстратом является глутаминовая кислота и равновесие реакции сдвинуто в сторону α-кетоглутарата
- синтез глутамина – взаимодействие глутамата с аммиаком. Является главным способом уборки аммиака, наиболее активно происходит в нервной и мышечной тканях, в почках, сетчатке глаза, печени. Реакция протекает в митохондриях.
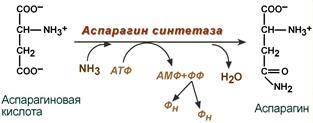
- синтез аспарагина – взаимодействие аспартата с аммиаком. Является второстепенным способом уборки аммиака, энергетически невыгоден, т.к. при этом тратятся 2 макроэргические связи,
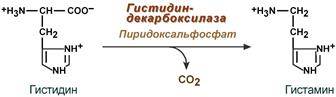
--- Декарбоксилирование аминокислот. Образование биогенных аминов и их роль в организме. Пути их распада. Синтез биогенных аминов (нейромедиаторов) из аминокислот связан с вовлечением в метаболизм α-карбоксильной группы аминокислот или, проще говоря, ее удалением. Гистамин Реакция образования гистамина наиболее активно идет в тучных клетках легких, кожи, печени, базофилах и эозинофилах. В них гистамин синтезируется и накапливается в секреторных гранулах.
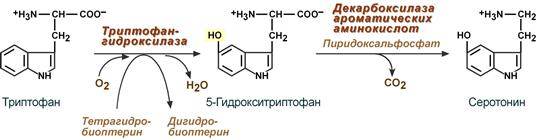
В кровь гистамин выделяется при повреждении ткани, при ударе, при электрическом раздражении. В клинической практике секреция гистамина обычно связана с аллергиями – при повторном попадании антигена в ранее сенсибилизированный организм развивается аллергическая реакция. Физиологические эффекты: - расширение артериол и капилляров и, как следствие, покраснение кожи, снижение артериального давления; - повышение проницаемости стенки капилляров и, как следствие, выход жидкости в межклеточное пространство (отечность), снижение артериального давления; - если предыдущие пункты имеют место в головном мозге – повышение внутричерепного давления; - увеличивает тонус гладких мышц бронхов, как следствие – спазм и удушье; - слабо повышает тонус мышц желудочно-кишечного тракта; - стимулирует секрецию слюны и желудочного сока. Серотонин Серотонин активно синтезируется в тучных клетках кожи, легких, печени, в селезенке, ЦНС.
Физиологические эффекты: - стимулирует сокращение гладких мышц желудочно-кишечного тракта и, как следствие, повышение перистальтики ЖКТ; - выражено стимулирует сокращение гладких мышц сосудов, кроме сосудов миокарда и скелетных мышц и, как следствие, повышение артериального давления; - слабо увеличивает тонус гладких мышц бронхов; - в центральной нервной системе является тормозным медиатором; - в периферических нервных окончаниях обусловливает возникновение боли и зуда (например, при укусе насекомых). Гамма-аминомасляная кислота Синтез γ-аминомасляной кислоты (ГАМК) происходит исключительно в центральной нервной системе в подкорковых образованиях головного мозга.
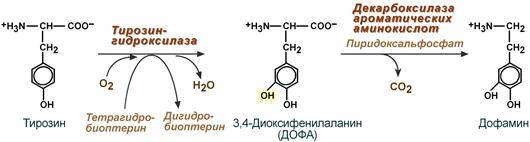
Физиологические эффекты В центральной нервной системе ГАМК (наряду с глутаминовой кислотой) является тормозным медиатором. Наиболее высока ее роль в височной и лобной коре, гиппокампе, миндалевидных и гипоталамических ядрах, черной субстанции, ядрах мозжечка. Дофамин Синтез дофамина происходит в основном в нейронах промежуточного и среднего мозга.
Физиологические эффекты Является медиатором дофаминовых рецепторов в подкорковых образованиях ЦНС, в больших дозах расширяет сосуды сердца, стимулирует частоту и силу сердечных сокращений, расширяет сосуды почек, увеличивая диурез. ---
|
|||||||
|
|
Последнее изменение этой страницы: 2021-05-12; просмотров: 714; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.238.20 (0.016 с.) |