
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
В гипоталамусе мужского организма «работает» только один центр, расположенный в среднем гипоталамусе. Полная блокада циклического центра преоптического гипоталаму-
ОБЩИЕ ЗАМЕЧАНИЯ Половые железы (гонады) вырабатывают половые гормоны. К ним относятся женские половые гормоны, в том числе эстрогены (эстрон, эстрадиол и эстриол), прогестины (в том числе — прогестерон), а также мужские половые гормоны — андрогены (в том числе — тестостерон). Все они являются гормонами стероидной природы, синтезируемыми из единого предшественника — холестерина. К половым гормонам следует также отнести гормональные факторы, которые участвуют в регуляции их продукции — гонадолиберин, фолли- тропин (ФСГ — фолликулостимулирующий гормон), лютропин (ЛГ—лютеинизирующий гормон). В рамках гормональной регуляции процессов размножения рассматривается также группа гормонов, вырабатываемых в плаценте, в том числе хорионический гонадотропин, хорионический соматомаммотропин (плацентарный лакгоген), а также гормон аденогипофиза—пролактин и факторы, регулирующие его продукцию — пролактостатин и про- лактолиберин. Кроме того, в последние годы обнаружены вещества, играющие определенную роль в регуляции репродуктивной функции человека, например, ингибин, релаксин, фактор, ингибирующий развитие Мюллерова протока и др. ГОНАДОЛИБЕРИН Считалось, что в гипоталамусе вырабатываются два рилизинг-фактора: фоллилиберин, стимулирующий выделение в гипофизе ФСГ, и люлиберин, стимулирующий выброс из аденогипофиза ЛГ. Оказалось, что оба этих либерина являются декапептидом: содержат 10 аминокислотных остатков и идентичны друг другу. Поэтому в последнее время говорят о наличии одного гормона, регулирующего выброс ФСГ и ЛГ — о гонадолнбернне. Гонадолиберинпродуцирующие нейроны гипоталамуса сконцентрированы в гипоталамусе женского организма в двух местах — в преоптической части гипоталамуса и в средней части в области аркуатного ядра. Нейроны преоптической области осуществляют циклический выброс гонад олиберина и регулируют половой цикл — развитие фолликула и желтого тела — за счет циклического выделения ФСГ и ЛГ. Этот центр функционирует с начала полового созревания, достигая «зрелости» к моменту его завершения. До полового созревания этот центр имеет высокую чувствительность к эстрогенам и поэтому не функционирует, т. е. заблокирован эстрогенами.
Нейроны, расположенные в области среднего гипоталамуса (аркуатное ядро), осуществляют постоянный выброс гонадолиберина. Это скопление нейронов получило название тонического центра выработки гонадолиберина. Предполагается, что секреция гонадолиберина осуществляется с интервалом 60—90 минут. Поэтому при недостаточности продукции гонадолиберина, сопровождающейся нарушением полового цикла у женщин, введение гонадолиберина с терапевтической целью осуществляется тоже пульсирующим способом, для этого используются специальные инжекторы с программным устройством. Показано, что введение гонадолиберина по обычной методике (например, 3 раза в день) не дает желаемого эффекта, в то время как импульсное введение вызывает восстановление менструального цикла у женщин. В гипоталамусе мужского организма «работает» только один центр, расположенный в среднем гипоталамусе. Полная блокада циклического центра преоптического гипоталаму- Физиология человека 161 са у лиц мужского пола происходит в период эмбрионального развития под влиянием высоких концентраций тестостерона. Очевидно, что закрепление дифференцировки гипоталамуса по мужскому типу происходит и в первый год жизни. Многие факторы влияют на выделение гонадолиберина. Так, известно, что опиатные пептиды (энкефалины, эндорфины, динорфины), нейротензин, гистамин, субстанция Р, холецистокинин, вазоактивный кишечный полипептид, а также мелатонин, половые гормоны — угнетают продукцию гонадолиберина. Различные эмоциональные воздействия могут приостановить выделение гонадолиберина. Не случайно, в период войн наступает нак называемая «аменорея» военного времени. Стало ясно, что норадренергические, дофаминергические и серотонинергические нейроны воздействуют на выброс гонадолиберина, однако направленность этих влияний пока до конца не ясна. Не исключено, что норадренергические нейроны повышают его. При действии гонадолиберина на ФСГ- и ЛГ-продуцируюшие клетки аденогипофиза происходит активация аденилатциклазы и повышение внутриклеточной концентрации цАМФ, что повышает активность протеинкиназ и как результат—увеличивается продукция соответственно ФСГ или ЛГ. Почему в одних случаях (например, в начале менструального цикла) гонадолиберин повышает главным образом продукцию ФСГ, а в последующем (перед овуляцией и в лютеияовой фазе менструального цикла) — в основном ЛГ? Объясняется это тем, что концентрация рецепторов для гонадолиберина в клетках, продуцирующих ФСГ и ЛГ — разная, и она регулируется эстрогенами. Роль половых гормонов — эстрогенов и прогестерона, а также тестостерона — велика в регуляции выброса гонадолиберина. Тонический центр гипоталамуса чувствителен к эстрогенам: высокие концентрации эстрогенов усиливают образование гонадолиберина, а низкие — тормозят. У мужчин в гипоталамусе имеется фермент, превращающий андрогены в эстрогены. Поэтому и у мужнин регуляция выброса гонадолиберина тоже осуществляется за счет эстрогенов.
При беременности, как и в лютеиновую фазу цикла, продукция гонадолиберина тормозится прогестероном, что временно задерживает выброс ФСГ. ФОЛЛИТРОПИН И ЛЮТРОПИН (ФСГИ ЛГ) Оба гонадотропных гормона продуцируются базофильными клетками аденогипофиза (дельта-базофилы). Каждый гормон содержит примерно 200 аминокислотных остатков и состоит из 2 субъединиц: альфа- и бета-. Альфа-субъединша предназначена для защиты гормонов от протеолитического действия ферментов, а бета-субьединица оказывает собственно физиологический эффект. У женщин фоллитропин, или фолликулостимулирующий гормон, вызывает рост и созревание фолликула — усиливает митоз фолликулярных клеток и синтез ДНК в ядрах клеток, повышает чувствительность яичников к действию лютропина и обеспечивает нормальную секрецию эстрогенов. Лютропин (ЛГ) у женщин способствует вместе с ФСГ росту фолликула, но главное — обеспечивает процесс овуляции и развитие желтого тела, т. е. продукцию прогестерона. У мужчин ФСГ в период полового созревания стимулирует рост и развитие гормонопродуцирующих интерстицальных клеток Лейдига, а в половозрелом возрасте стимулирует сперматогенез и поддерживает развитие и функцию клеток Сертоли, которые необходимы для поддержания нормальных условий сперматогенеза. ЛГ (лютропин) стимулирует образование в клетках Лейдига андрогенов, в том числе тестостерона. ПОЛОВОЕ РАЗВИТИЕ, ИЛИ ПОЛОВАЯ ДИФФЕРЕНЦИРОВКА ГОНАД И ПОЛОВЫХ ОРГАНОВ Половая дифференцировка охватывает ряд этапов эмбрионального и постэмбрионального периодов, а понятие «пол» слагается из ряда взаимосвязанных компонентов — биологических, психологических и социальных. В данном разделе в основном представлены данные, касающиеся биологического компонента. Знание этих вопросов является ключом к пониманию патологии и отклонений в сексуальном поведении человека. Генетический пол будущего ребенка предопределяется в момент слияния яйцеклетки и сперматозоида. Он обусловлен набором половых хромосом, образующихся в зиготе при соединении материнских и отцовских гамет. Кариотип 46 XX дает развитие по женскому типу, а кариотип 46 XY — по мужскому. Однако путь к мужскому организму достаточно сложен — масса всевозможных условий должна быть налицо, чтобы произошло становле- ниемужскогоорганизма.
До 6 недель внутриутробного развития мужские и женские половые железы развиваются из одного недифференцированного зачатка, который состоит из коркового и мозгового вещества (корковое вещество в последующем переходит в яичник, а мозговое — в яичко). Имеется также Мюллеров проток, который в женском организме переходит в матку и маточные трубы, и Вольфов проток, на основе которого развиваются в мужском организме внутренние половые органы. Ген, детерминирующий дифференцировку зачатка гонады по мужскому типу, содержится в Y-хромосоме. Благодаря этому гену в организме начинает синтезироваться специфический мембранный белок, получивший название H-Y-антигена. В клетках развивающегося организма, в том числе в клетках гонады содержится рецептор к этому белку. Взаимодействие H-Y-антигена с этими рецепторами индуцирует превращение первичной гонады в яичко. Если самке в период эмбриального развития ввести H-Y-антиген, то ее развитие тоже пойдет по мужскому типу, хотя кариотип у нее женский. В последние годы показано, что, вероятно, существует несколько вариантов антигена типа H-Y и соответственно несколько генов (до 18), контролирующих их синтез. Возможно, что для дифференцировки первичной гонады в яичник необходимы тоже специфические факторы, подобные H-Y-антигену, которые, вероятно, контролируются генами Х-хромосомы. Дифференцировка гонады в яичник заключается в том, что гоноциты дифференцируются в овогонии, а мезенхимальные клетки дифференцируются в тека-ткань; дифференцировка гонады в яичко проявляется в том, что гоноциты дифференцируются в сперматогонии, а интерстициальныеклетки — в клетки Лейдига. ______ В эмбриональном периоде яичник не продуцирует половые гормоны, а яичко обладает выраженным андрогенпродуцирующим свойством. Благодаря такой разнице в эмбриональном периоде происходит дифференцировка внутренних и наружных половых органов. У женского организма этот процесс дифференцировки не нуждается в продукции специальных гормонов — дифференцировка предопределена генетически: в результате из Мюллерова протока образуется матка и маточные трубы (матка образуется за счет слияния Мюллеро- вых протоков на 9—11 нед.). Наружные половые органы дифференцируются у женщин с 12 по 20 нед. внутриутробного периода. Они развиваются из урогенитального синуса, полового бугорка и лабиосакральных валиков: из них формируются независимо от состояния гонады влагалище, клитор, большие и малые половые губы, преддверие влагалища.
У организма мужского пола дифференцировка внутренних половых органов (придатка яичка, семявыносяшего протока, семенных пузырьков) и наружных половых органов (полового члена, мошонки, предстательной железы и мочеиспускательного канала) происходит при обязательном участии андрогенов, в том числе — тестостерона (из Вольфова протока образуются придатки яичка, семявыносяший проток, семенные пузырьки, из урогенитального синуса — предстательная железа и мочеиспускательный канал, из полового бугорка ■— половой член и кавернозные тела, из половых валиков — мошонка). Каким же образом происходит дифференцировка в мужском организме? Основной фактор — это тестостерон. Кроме того, в клетках Сертоли вырабатывается фактор, который вызывает регрессию Мюллерова протока. Этот фактор получил название антимюллеров- ский фактор, мюллероподавляющая субстанция. Он представляет собой гликопротеин. Те- 163 стостерон как основной андрогенный гормон продуцируется в клетках Лейдйга. Эти клетки начинают функционировать в яичке, начиная с 8—12 недели внутриутробного развития, под влиянием материнского хорионического гонадотропина, а в последующем (до конца внутриутробного развития) — под влиянием ФСГ и ЛГ, в том числе и ЛГ плода. Итак, под влиянием прежде всего хорионического гонадотропина в крови плода резко возрастает концентрация тестостерона, которая достигает уровня, характерного для половозрелого мужчины. Тестостерон воздействует на клетки-мишени, в которых под влиянием фермента 5-альфа-редуктаэы он превращается в собственно гормон 5-альфа-дигидротестостерон. Именно в такой форме он может оказывать свой физиологический эффект — влиять на дифференцировку клеток-мишеней. В клетке дигидротестостерон соединяется со специфическими рецепторами, достигает в таком виде ядра, связывается в нем с хроматином, в результате — активируются соответствующие гены и стимулируется синтез необходимых белков. Рецепторы, с которыми связывается дигидротестостерон, регулируются геном, расположенным в Y-хромоеоме. Помимо влияния на развитие внутренних и наружных половых органов, тестостерон влияет на дифференцировку гипоталамуса по мужскому типу —" блокирует развитие циклического центра продукции гонадолиберина. Итак, для того, чтобы осуществилась дифференцировка организма по мужскому типу, необходимо, чтобы происходили: 1) секреция тестостерона клетками Лейдйга, 2) секреция клетками Сертоли фактора, угнетающего развитие протока Мюллера, 3) наличие в клетках-мишенях 5-альфа-редуктазы, 4) наличие в клетках-мишенях цитоплазматических рецепторов к дигидротестостерону. Любой дефицит в этих звеньях, любое нарушение приводит к дефекту половой дифференцировки. ЖЕНСКИЕ ПОЛОВЫЕ ГОРМОНЫ. ЭСТРОГЕНЫ. ПРОГЕСТИНЫ
Вырабатываются в надпочечниках, яичнике, а во время беременности—в фетоплацентар- ной системе (в плаценте продуцируются предшественники эстрогенов, а плод завершает их синтез, поэтому когда уровень эстрогенов низкий, то это свидетельствует о фетоплацентар- ной недостаточности). В организме имеется три вида эстрогенов: эстрон, эстрадиол и эстриол (соответственно, их обозначают как Э„ Э2 и Э3 — по количеству гидроксильных групп). Наибольшей физиологической активностью обладает эстрадиол. Эстриол является метаболитом эстрадиола и эстрона и в физиологическом плане мало активен. В крови эстрогены связываются глобулинами, синтезируемыми в печени, эти же глобулины связывают и андрогены. Метаболизм эстрогенов происходит в печени. Здесь они и их метаболиты связываются с глюкуроновой и серной кислотами и экскретируются из организма с желчью и мочой. Действуют эстрогены путем изменения синтеза белков в клетке, хотя могут оказывать и внерецепторный эффект. Физиологические эффекты эстрогенов: 1) Угнетают секрецию ФСГ и ЛГ, снижают ответ аденогипофиза на гонадолиберин и таким образом участвуют в регуляции созревания и развития фолликула. 2) Стимулируют рост и развитие внутренних и наружных половых органов. Недостаточ ность эстрогенов приводит к недоразвитию этих органов, инфантилизму. 3) Стимулируют развитие вторичных половых признаков. 4) Усиливают процессы кровообращения в половых органах. 5) Приводят к изменению слизистой матки — вызывают процесс пролиферации эндоме трия. 6) Обладают выраженным анаболическим эффектом. 7) Усиливают обмен костной ткани, ускоряют созревание скелета (поэтому при раннем половом созревании быстро прекращается эпифизарный рост костей и рост в длину оста навливается). 8) В больших дозах ведут себя как минералокортикоиды (альдостерон): вызывают за держку в организме натрия и воды. Возможно, что это имеет место при беременности, что выражается в развитии нефропатии. А
ДЕПОНИРОВАНИЕ ЖИРА |
| |||||||||||||||||||
|
| ПОВЫШЕНИЕ t° ТЕЛА. СИНЕРГИЗМ С АНДРОГЕНАМИ |
| |||||||||||||||||||
|
|
| ||||||||||||||||||||
| ТОРМОЖЕНИЕ ТИМУСА | It |
| ПРИ БЕРЕМЕННОСТИ: ОБЕСПЕЧЕНИЕ ИМПЛАНТАЦИИ ПЛОДА | ||||||||||||||||||
|
| |||||||||||||||||||||
|
СТИМУЛЯЦИЯ СЕКРЕЦИИ АКТГ |
|
| |||||||||||||||||||
| РАЗВИТИЕ ЛАКТАЦИИ |
Б рТ| Выделение ФСГ
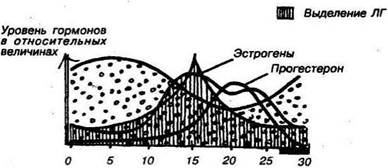
Рис. 49. Женские половые гормоны (эстрогены и прогестерон) и их' функции (А).
Дни менструального цикла и преобладание различных гормонов на протяжении определенных его фаз (Б). ФСГ — фолликулостимулирующий гормон. ЛГ — лютеинизирующии гормон.
Особую роль играют эстрогены при беременности. В результате -Высокого содержания всех фракции эстрогенов при беременности происходит значительный рост мышечной ткани матки (масса матки возрастает с 50 г до 1 кг) за счет гиперплазии и гипертрофии. Эстрогены способствуют эффективному маточно-плацентарному кровообращению, а также развитию необходимых для процесса плодоизгнания мышечных плазматических рецепторов: ОКСИТОЦИНОВЫХ СерИТИНИНИНЫХ, ГНСтамИНОВЫХ. ВмеСТС С нрОгеСГСрОИОМ Н ирОЛаКТНИОМ эстрогены способствуют развитию молочных желез. Накануне родов эстрогены причастны к индукции родового акта.
Прогестины. Это гормоны, необходимые для развития беременности. Один из них — прогестерон. Он продуцируется в коре надпочечников, но основное место его образования — это желтое тело, а во время беременности после 12—16 недель — плацента. В крови
165 связывается транспорта ном. Метаболизируется в печени, где подобно эстрогенам связывается глюкуроновой и серной кислотами и выводится с мочой.
Его функции многообразны и до конца не установлены. У небеременных женщин прогестерон участвует в регуляции полового цикла: выделяясь в лютеиновую фазу цикла, он тормозит продукцию ФСГ и тем самым пролонгирует половой цикл. Прогестерон способствует усилению секреции железами эндометрия, в результате чего в секрете возрастает концентрация гликогена, необходимого для развития эмбриона. Прогестерон обладает рядом важных физиологических эффектов — в том числе задерживает в организме натрии и воду, а также оказывает выраженный пирогенный эффект — повышает температуру тела на 0,4—0,5 град. Этот эффект широко используется в медицинской практике для определения времени наступления овуляции, так как после овуляции начинает функционировать желтое телои продуцироватьсяпрогестерон.
Во время беременности прогестерон вместе с эстрогенами способствует морфологическим перестройкам в матке, миометрии, молочных железах. Считалось, что прогестерон тормозит сократительную деятельность матки и блокирует эффекты стимуляторов, способствует вынашиванию плода. Широкое распространение получила в 50—60-е годы теория прогестеронового блока Чапо. В опытах на животных показано, что роды не могут начаться, если уровень продукции прогестерона не снизится до минимальных значений. Это типично для кролика, крысы, свиньи. У женщин, однако, ситуация иная. Накануне родов уровень прогестерона не только не снижается, а наоборот, имеет тенденцию к повышению. Следовательно, роды у женщин проходят на фоне высокого содержания прогестерона. Значит, прогестерон не способен тормозить сократительную деятельность матки у женщин? Вероятно, это положение справедливо для 2-го и 3-го триместров беременности, когда на помощь прогестероновому механизму приходит бета-адренорецепторный ингибирующий механизм, вызывающий угнетение спонтанной и вызванной сократительной активности миометрия. В первом триместре прогестерон, возможно, все-таки оказывает прямое тормозное влияние на матку: при его недостаточности имеет место угроза прерывания беременности или выкидыш.
Учитывая, что во время родов происходит значительное повреждение миометрия, которое, например, может приводить к полной утрате миометрием чувствительности к окситоцину, серотонину и другим утеростимуляторам, выдвинуто предположение о том, что прогестерон играет роль фактора, препятствующего повреждению миометрия. Однако пока прямых доказательств правильности этой гипотезы не получено.
РАЗВИТИЕ ФОЛЛИКУЛА. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ПОЛОВОГО ЦИКЛА
При дифференцировке гонады по женскому типу из коркового слоя первичной гонады образуется яичник. В нем находятся примордиальные фолликулы. Каждый фолликул содержит яйцеклетку (овоиит), окруженную одним рядом клеток фолликулярного эпителия. Рост примордиального фолликула выражается в увеличении рядов клеток фолликулярного эпителия, в результате чего формируется зернистая оболочка (зона гранулеза).
Первоначальные стадии роста фолликула (до 4 слоев эпителиальных клеток) происходят автономно, без участия ФСГ и ЛГ. Для дальнейшего созревания требуется ФСГ — под его влиянием растет число слоев зернистой оболочки. Но это происходит уже в период половой зрелости. К моменту рождения девочки число примордиальных фолликулов достигает 300—400 тыс.
Под влиянием повышения выброса в кровь ФСГ происходит рост числа слоев зернистого эпителия. При этом эпителий начинает вырабатывать жидкость, которая формирует полость фолликула. Одновременно под влиянием ФСГ зернистый эпителий начинает вырабатывать эстрогены. На этой стадии развития фолликул носит название граафова пузырька, или пузырчатого фолликула.
Вокруг граафова пузырька имеются клетки стромы. Они формируют две оболочки: внутреннюю и внешнюю (тека-интерна и тека-экстерна). Клетки внутренней оболочки (тека- клетки внутреннего слоя) продуцируют эстрогены, а клетки теки-наружной — андрогены.
Процесс очередного созревания фолликула в период половой зрелости начинается с воздействия ФСГ на те фолликулы, которые имеют рецепторы к ФСГ. Обычно это 10—IS фолликулов, но созревание наступает лишь для одного фолликула, остальные подвергаются атрезии за счет внутрияичникового механизма саморегуляции. Под влиянием ФСГ, как уже отмечалось выше, растет число клеток фолликулярного эпителия, увеличивается объем фолликула и повышается продукция эстрогенов. Максимальная концентрация ФСГ в крови достигается к 7—9 дню от начала созревания фолликула (цикла). Продукция ЛГ тоже осуществляется под влиянием гонадолиберина, но увеличение концентрации ЛГ в крови идет меньшими темпами и с меньшей скоростью, чем рост уровня ФСГ. За счет повышения продукции эстрогенов происходит снижение продукции ФСГ (но не ЛГ). Когда концентрация эстрогенов достигает максимального значения, это вызывает активацию гонадолиберинпро- дуцируюших нейронов циклического центра гипоталамуса (преоптическая область) и повышение выброса гонадолиберина. Это в свою очередь приводит к дополнительному выбросу ФСГ и, что особенно важно, ЛГ. Значительно возрастает продукция фолликулярной жидкости и происходит разрыв граафова пузырька—выход яйцеклетки из яичника в брюшную полость, откуда она попадает в маточные трубы и в матку. Обычно пик ЛГ происходит за 24 часа до овуляции. На месте разорвавшегося фолликула образуется желтое тело — клетки зернистого слоя гипертрофируются, накапливают желтый пигмент лютеин и начинают продуцировать прогестерон. Благодаря прогестерону продукция ФСГ тормозится, а продукция ЛГ сохраняется на высоком уровне, что важно для функционирования желтого тела. Одновременно начинает возрастать уровень пролактина (за счет выделения пролакто- либерина гипоталамуса), что сопровождается торможением продукции ФСГ. Таким образом, прогестерон и пролактин способствуют тому, чтобы уровень ФСГ в этот период не возрастал и не приводил к созреванию новой партии фолликулов.
В случае, если происходит оплодотворение яйцеклетки, то хорионический гонадотропин, продуцируемый с первых же часов существования оплодотворенной яйцеклетки, а в последующем — и плацентарный лактоген поддерживают продукцию прогестерона в желтом теле. После формирования плаценты роль желтого тела снижается, но все-таки до конца беременности оно продолжает функционировать как источник прогестерона.
При отсутствии сигнала от яйцеклетки (отсутствии хорионического гонадотропина) происходит редукция желтого тела со снижением продукции прогестерона. Вероятно, этот процесс происходит под влиянием простагландинов, уровень которых к концу лютеиновой фазы цикла возрастает и приводит к изменению яичникового кровотока, что и вызывает лютеолиз — редукцию желтого тела. Одновременно простагландины активируют продукцию ФСГ, и на фоне уменьшения уровня прогестерона и пролактина происходит усиление выделения из тонического центра гипоталамуса гонадолиберина, что усиливает продукцию ФСГ и ЛГ. Цикл вновь повторяется.
Принято различать яичниковый цикл и маточный цикл.
У женщин весь менструальный цикл в среднем продолжается 28 дней. У части женщин — 21,26 или 32 дня. При 28-дневном цикле яичниковый цикл состоит из трех фаз, каждая из которых занимает следующий промежуток времени:
1) фолликулярная фаза — с 1 по 14 день цикла;
2) овуляторная фаза или фаза овуляции — 14-й день цикла;
3) лютеиновая фаза — с 15 по 28 день.
Соответственно, уровень половых гормонов меняется так: в фолликулярную фазу цикла постепенно возрастает уровень эстрогенов, достигая максимума к моменту овуляции (а точнее — за сутки до овуляции), ас 15 дня — с начала лютеиновой фазы — преобладает уровень прогестерона.
В матке, в ее эндометрии в период цикла происходят определенные изменения. Их делят на 4 фазы: десквамации, регенерации, пролиферации и секреции.
Фаза десквамации представляет собой отторжение эндометрия — менструацию. Она (фаза) продолжается 3—5 дней. Почти параллельно с ней начинается фаза регенерации —
167 восстановления эндометрия, которая завершается к 5—6 дню от первого дня менструации. Затем наступает фаза пролиферации — рост эндометрия, развитие в нем желез. Фаза длится до 14 дня (до момента овуляции). Фаза секреции — от 15 до 28 дня включительно — под влиянием прогестерона железы эндометрия начинают продуцировать секрет, содержащий гликоген. Таким образом, в этой фазе слизистая матки готова принять для дальнейшего развития оплодотворенную яйцеклетку. Затем за счет сужения сосудов происходит некроз и начинается вновь фаза десквамации (отторжения эндометрия).
При нарушении гормональной регуляции (гипоталамические, гипофизарные, яичниковые нарушения) у женщин может наблюдаться аменорея — отсутствие менструаций, отсутствие развития фолликула, или наблюдаются циклические нарушения, при которых менструальные циклы есть, но они нарушены (разное количество крови выделяется в момент менструации). Наконец, существуют так называемые ановуляторные циклы, при которых фолликул зреет, но овуляция не возникает, в результате чего происходит атрезия данного фолликула и все повторяется заново. В этом случае имеет место лишь одна фаза (в яичнике
— фолликулярная, в матке — фаза пролиферации), а менструальноподобные кровотечения происходят в период регрессии фолликула.
Примечания. 1. В некоторых учебниках дается иная классификация женского полового цикла. Так, по Г. И. Косицкому, весь половой цикл разделен на 4 фазы: предовуляционный, овуляционный, послеовуляционный (метаэструс) и период покоя (диэструс). Предовуляционный период (проэструс) — это период созревания фолликула (соответствует фолликулярной фазе яичникового цикла), овуляционный — это период от момента овуляции до менструации (соответствует овуляторной и лютеиновой фазам приведенной выше классификации). Послеовуляционный период (метаэструс) — соответствует моменту появления менструации. Наконец, завершающий этап — период покоя — от момента последней менструации до начала созревания фолликула. Таким образом, эта классификация не соответствует классификации, принятой в акушерстве и гинекологии.
2. У животных, не относящихся к приматам (крыса, кролик, свинья, корова и т. п.) отсутствуют менструации. У них используется следующая классификация фаз полового цикла. 1) Фаза предтечки (проэструс) — период созревания фолликула до овуляции. 2) Фаза течки
— период овуляции, который сопровождается влагалищным кровотечением и особым пове дением: возникает состояние «охоты», или течки, состояние, при котором самка «принима ет» самца. 3) Фаза послетечки (метаэструса) и фаза межтечки (диэструса). Например, у лабораторных крыс весь половой цикл (астральный цикл) занимает около 4 суток, и каждая фаза длится примерно 1 сутки.
Диагностика фаз цикла. Перед акушером-гинекологом часто возникает задача: определить, происходит ли у женщины овуляция, имеет ли место нормальное течение полового цикла. Одним из способов диагностики правильности течения менструального цикла является определение уровня гормонов: ФСГ, ЛГ, эстрогенов, прогестерона. Но для этого требуются соответствующая аппаратура и условия. В клинической практике используются более доступные методы. Например, определение базальноя (ректальной) температуры: по утрам у женщины проводится измерение температуры. С момента овуляции ив последующие дни менструального цикла, т. е. в период лютеиновой фазы яичникового цикла температура на 0,4—0,5 градуса выше, чем в период фолликулярной фазы. Таким образом, определение базальной температуры позволяет достаточно просто и точно определить динамику полового цикла. Второй способ — это определение картины влагалищного мазка — наличие изменений в эпителии указывает на нормальное течение полового цикла женщины.
У животных, в том числе у крысы, мыши также используется методика' определения картины влагалищного мазка. Так, например, в фазу проэструса (предтечки) влагалищный мазок состоит почти исключительно из округлых и многоугольных эпителиальных клеток с зернистою и довольно крупной цитоплазмой. В фазе эструса (течки) влагалищный мазок состоит только из крупных ороговевших безъядерных клеток, имеющих вид чешуек неправильной формы. В конце течки ороговевшие клетки образуют скопления. В фазе послетеч- ки влагалищный мазок содержит много ороговевших чешуек и лейкоцитов. В фазу межтечки (диэструса) влагалищный мазок содержит значительное количество слизи, много лейкоцитов и небольшое число эпителиальных клеток с ядрами.
Итак, влагалищный мазок может служить показателем эндокринной функции яичников, гипофиза и гипоталамуса.
МУЖСКИЕ ПОЛОВЫЕ ГОРМОНЫ. АНДРОГЕНЫ
Мужские половые гормоны вырабатываются у мужчин и женщин в надпочечниках (сетчатая зона коры), в яичках мужчин и в яичнике (наружный тека-слой) женщин. Основное место продукции андрогенов — яичко.
Яичко представляет собой мужскую половую гонаду, в которой происходит сперматогенез и продукция андрогенов. Сперматогенез осуществляется в извитых канальцах, которые выстланы первичными половыми клетками — сперматогониями и клетками Сертоли. Клетки Сертоли питают сперматозоиды. Андрогены продуцируются клетками Лейдига — это интерстициальные клетки яичка, совокупность этих клеток часто называют пубертатной железой. Синтезируются андрогены из холестерина, главным образом под влиянием ФСГ и ЛГ. Существуют два варианта синтеза андрогенов (холестерин -> прегненолон -> прогестерон -> 17-альфа-гидроксипрогестерон -> андростендион -> тестостерон (1-й вариант) и прегненолон -> 17-альфа-гидроксипрегненолон -> дегидроэпиандростерон -> андростендион -> тестостерон -> (2-й вариант). Андростерон, дегидроэпиандростерон и тестостерон являются андрогенами (андрос — мужчина), но наиболее активным является тестостерон. В крови андрогены связываются глобулином, который синтезируется в печени и который одновременно способен переносить и эстрогены. 98% тестостерона находится в крови в связанном состоянии. Когда уровень белка возрастает (это, к примеру, имеет место при циррозе печени, гипертиреозе, гипонанизме), то возрастает и доля связанного с белками тестостерона. Активным же является свободный тестостерон. За сутки у мужчин синтезируется 6—7 мг тестостерона и его концентрация в крови достигает 2,7—8,5 нмоль/л. Для сравнения: у женщин — соответственно 1 мг/сутки и 0,6—1,9 нмоль/л.
Тестостерон разрушается в печени под влиянием 17-бета-дегидрогеназы, а его метаболиты связываются с глюкуроновой и серной кислотами и экскретируются с мочой в виде 17-кетостероидов.
Уровень тестостерона в крови мужского организма достигает высоких значений в эмбриональном периоде, в первые 3 месяца после рождения, затем снижается, небольшой подъем наблюдается после 6 лет (в период препубертата), но значительно начинает возрастать в период полового созревания и снижается после 50 лет. Уровень тестостерона имеет суточные колебания: максимальные значения наблюдаются в 7—9 час. утра, а минимальные — в 24—3 час.
Механизм действия тестостерона связан с тем, что в клетках-мишенях происходит его превращение: в нейронах гипоталамуса, продуцирующих гонадолиберин (тонический центр в среднем гипоталамусе), тестостерон превращается в эстроген, который оказывает тормозное действие на продукцию гонадолиберина. Тем самым поддерживается уровень тестостерона на определенных значениях. В других клетках-мишенях — в том числе в клетках Сертоли, во внутренних и наружных половых органах, в других тканях — тестостерон с помощью 5-альфа-редуктазы превращается в 5-альфа-дегидротестостерон, который входит внутрь цитоплазмы клетки-мишени, связывается там с цитоплазматическими рецепторами, образованный комплекс входит в ядро, где осуществляется активация транскрипции м-РНК и р-РНК (активируется синтез соответствующих белков).
Физиологические эффекты тестостерона:
— участвует в половой дифферешдировке гонады и в развитии первичных половых признаков (внутренних и наружных половых органов), в развитии вторичных половых признаков (оволосение и организация скелета по мужскому типу);
■— стимулирует рост скелета и всех тканей организма, ускоряет созревание скелета, прекращает эпифизарный рост трубчатых костей, чем ограничивает рост организма в длину;
169
— обладает анаболическим эффектом — стимулирует белковый синтез в скелетной му скулатуре, в других тканях;
— обеспечивает либидо и потенцию, обеспечивает мужской тип полового поведения;
— участвует в регуляции сперматогенеза (за счет воздействия на клетки Сертоли).
Избыточное содержание тестостерона вызывает гиперсексуальность, повышенный рост волос.
Регуляция уровня тестостерона осуществляется за счет его прямого влияния на гипоталамус (гонадолиберинпродуцирующие нейроны тонического центра) -> выделение гонадолиберина -> выделение ЛГ -> продукция тестостерона в яичке клетками Лейдига. Когда уровень тестостерона повышен, продукция гонадолиберина снижается и наоборот.
ПРОЛАКТИН, ПРОЛАКТОСГАТИН, ПРОЛАКТОЛИБЕРИН
Пролактин продуцируется в аденогипофизе. Это полипептид, содержащий 198 аминокислотных остатков. Пролактин, СТГ и хорионический соматомаммотропин (плацентарный лактоген) имеют много общего по своему строению. Возможно, они происходят из одного и того же источника.
Физиологические эффекты пролактина:
—участвует в регуляции осмотического давления (за счет предупреждения избыточной потери ионов натрия и воды). Очевидно, это имеет значение при беременности, когда уровень пролактина возрастает (как и эстрогенов), что вызывает задержку ионов натрия и воды в организме;__________________
— вызывает анаболический эффект (повышает синтез белков);
— у женщин пролактин участвует в регуляции менструального цикла, его уровень возра стает в лютеиновую фазу цикла, и тем самым гормон способствует сохранению желтого тела и продукции прогестерона. Одновременно пролактин тормозит продукцию гонадолиберина и ФСГ, под влиянием пролактина тормозится созревание очередного фолликула;



