Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Роль печени в поддержании нормального уровня содержания глюкозы в крови.
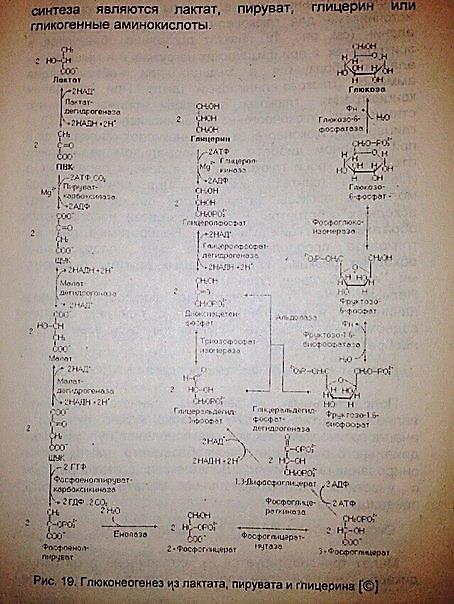
Гликоген печени используется для поддержания относительного постоянства концентрации глюкозы в крови. Печень обладает способностью реагировать на изменение концентрации сахара в крови. Обладая местным гомеостатическим механизмом, печень при уменьшении концентрации сахара в крови выделяет большее количество его за счет гликогенолиза, а при увеличении — меньшее; при значительной же гипергликемии печень использует глюкозу для синтеза гликогена и липидов. 35.. Глюконеогенез: начальные субстраты, клеточная локализация, реакции, ферменты и коферменты, биоэнергетика, регуляция, значение для организма (рассмотреть на примере глюконеогенеза из лактата). Глюконеогенез -это образование глюкозы из таких неуглеводных предшественников,как лактата,промеж.продукты циклокарбон. К-т,глицерола,пропиата. Процесс протекает в основном в печени и менее интенсивно в кор-ковом веществе почек, а также в слизистой оболочке кишечника. Первичными субстратами глюконеогенеза являются лактат, аминокислоты и глицерол . Лактат является продуктом анаэробного гликолиза. Он образуется прилюбых состояниях организма (эритроциты, работающие мышцы). Таким образом, лактат используется в глюконеогенезе постоянно. • Глицерол высвобождается при гидролизе жиров в жировой ткани в период голодания или при длительной физической нагрузке. • Аминокислоты образуются в результате распада мышечных белков ивключаются в глюконеогенез при длительном голодании или продолжительной мышечной работе. Схема включения субстратов.
Цикл Кори обеспечивает утилизацию лактата, предотвращает накопление лактата и,как следствие этого, лактоацидоз(опасное снижение pH,когда равновесие в реакции пируват-лактат сдвигается в сторону лактата. Реакции: Необратимые: 1,4,11,13!!!! 1)2ПВК+2АТФ+2СО2->(пируваткарбоксилаза)2 оксалоацетат+2фосфат неорганич.+2 АДФ 2)2 ацелоацетат+ 2 НАДН(Н+)->(малатдегидрогеназа) 2 малат+2НАД+ 3)2 малат +2НАД+->(малатдегидрогеназа)2 ацелоацетат+2НАДН(Н+) 4)2 оксалоацетат+2 ГТФ->(фосфоенол-пируват-карбоксикиназа)2 фосфоенол-пируват+2ГДФ 5)2 фосфоенол-пируват->(енолаза)2 фосфоглицерат 6)2 фосфоглицерат->(фосфоглицератмутаза) 2 3-фосфоглицерат 7)2 3-фосфоглицерат +2АТФ->(фосфоглицераткиназа)2 1,3-дифосфоглицерат +2 АДФ
8)2 1,3-дифосфоглицерат +2 НАДН(Н+)->(глицеральдегид-3-фосфат-дегидрогеназа) 2 глицеральдегид-3-фосфат+2 фосфат нерганич. 9)Глицеральдегид-3-фосфат->(триозофосфат-изомераза) дигидрокси ацетон фосфат 10)Глицеральдегид-3-фосфат+дигидрокси-ацетон-фосфат->(фруктозодифосфат-альдалаза) фруктозо-1,6-дифосфат 11)Фруктозо-1,6-дифосфат+Н2О->(фруктозодифосфатаза) фруктозо-6-фосфат+фосфат неорганич. 12)Фруктозо-6-фосфат->(фосфоглико-изомераза) глюкозо-6-фосфат 13)Глюкозо-6-фосфат + Н2О->(глюкозо-6-фосфатаза) глюкоза+ фосфат неорганич. Регуляция глюконеогенеза: Превращение метаболитов в глюконеогенезе связаны так,что один процесс активен,а второй неактивен. А вообще термодинамически выгодна их одновременная активация. 1.Ацетил-КоАактивирует,а АДФ ингибирует пируваткарбоксилазу(первый фермент глюконеогенеза). Когда в клетке накапливается митохондриальный ацетил-КоА,биосинтез глюкозы из пируватаусиливается.,одновременно подавляются окислительные декарбоксилирования пирувата. Скорость глюконеогенеза зависит от концентрации лактата и других предшественников глюкозы 2. Фруктоза-1,6-бисфосфотаза ингибируется АМФ. В тоже время АМФ является аллостерическим активатором фосфофруктокиназы. При низкой концентрации АМФ и высокой концентрации АТФ происходит стимуляция глюконеогенеза. А когда величина АМФ низкая,в клетке наблюдается расщепление глюкозы. Фруктозо-2,6-бисфосфатаза активирует фосфо-фруктокиназу и ингибирует фруктозодифосфатазу Роль глюконеогенеза в обмене углеводов: глюконеогенез не только обеспечивает органы и ткани глюкозой, но еще и перерабатывает образующийся в тканях лактат, препятствуя тем самым развитию лактат-ацидоза.
36. Глюконеогенез из глицерина. Схема в 35 вопросе 37. Глюконеогенез из пировиноградной кислоты.
Первая необратимая реакция глюконеогенеза катализируется митохондриальной пируваткарбоксилазой, которая содержит в качестве кофермента витамин Н (биотин). В митохондриях этот фермент катализирует АТФ-зависимуюреакцию карбоксилирования пирувата, в ходе которой образуется оксалоацетат. Для оксалоацетата внутренняя мембрана митохондрий непроницаема, и транспорт его в цитоплазму происходит с помощью малатного челночного механизма.Митохондриальная малатдегидрогеназа восстанавливает оксалоацетат до малата, который может выходить в цитоплазму. Затем уже цитоплазматическая малатдегидрогеназа окисляет малат до оксалоацетата для последующего участия в реакции
После того как в мыщцах истощается запас гликогена, основным источникомпирувата становятся аминокислоты, образующиеся после деградации белков. При этом более 30% аминокислот, поступающих из крови в печень, приходится на аланин — одну из гликогенных аминокислот, углеродный скелет которой используется в печени как предшественник для синтеза глюкозы. Механизм превращения мышечных аминокислот в аланин, схема его участия в глюконеогенезе представлены в гл. 24. Другим источником пирувата является лактат, который накапливается в интенсивно работающих мышцах в процессе анаэробного гликолиза, когда митохондрии не успевают реокислить накапливающийся НАДН. Лактат транспортируется в печень, где снова превращается в пируват, а затем в глюкозу и гликоген. Схема в 35 вопросе
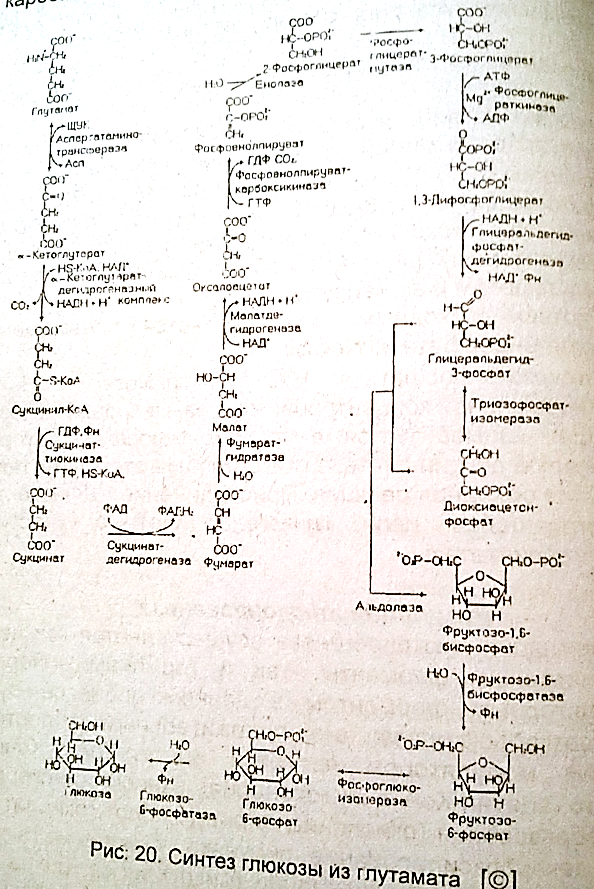
38.. Глюконеогенез из глутаминовой кислоты.
39. Пентозофосфатный цикл: локализация процесса, реакции, ферменты и коферменты, биоэнергетика, регуляция, значение для организма … представляет собой прямое окисление глюкозы и протекает в цитоплазме клеток. Наибольшая активность ферментов пентозофосфатного пути обнаружена в клетках печени, жировой ткани, коры надпочечников, молочной железы в период лактации, зрелых эритроцитах. Низкий уровень этого процесса выявлен в скелетных и сердечной мышцах, мозге, щитовидной железе, легких. Пентозофосфатный путь выполняет в организме две важнейшие метаболические функции: он является главным источником НАДФН для синтеза жирных кислот, холестерола, стероидных гормонов, микросомального окисления; в эритроцитах НАДФН используется для восстановления глутатиона – вещества, препятствующего пероксидному гемолизу; он является главным источником пентоз для синтеза нуклеотидов, нуклеиновых кислот, коферментов (АТФ, НАД, НАДФ, КоА-SН и др.).
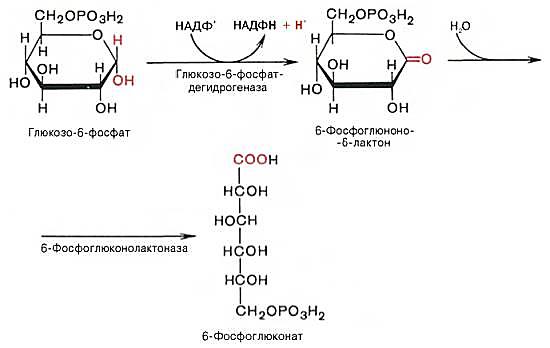
Первой реакцией пентозофосфатного пути является ферментное дегидрирование глюкозо-6-фосфата в глюконолактон-6-фосфат, протекающее с участием НАДФ+ в качестве кофермента. Далее 6-фосфоглюконолактон гидратируется и превращается в 6-фосфоглюконовую кислоту
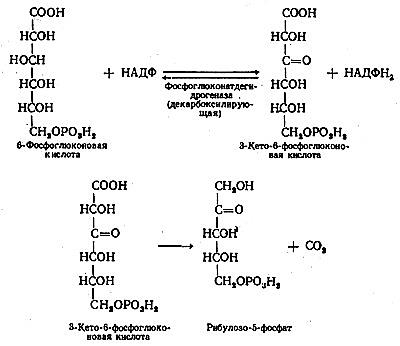
На следующей стадии 6-фосфоглюконат подвергается дегидрированию с участием НАДФ+-6-осфоглюконатдегидрогеназы и образованием 3-кето-6-фосфоглюконовой кислоты и НАДФН(Н+) Образовавшаяся 3-кето-6-фосфоглюконовая кислота декарбоксилируется с образованием D-рибулозо-5-фосфата и СО2. 6 молекул рибулозо-5-фосфата изомеризуется в 4 молекулы ксилулозо-5-фосфата и 2 молекулы рибозо-5-фосфата. В случае высокой потребности в рибозо-5-фосфата реакции пентозофосфатного пути заканчиваются образованием этих метаболитов, а при высокой потребности в НАДФН(Н+) – продолжается далее.
Рибулозо-5-фосфат при участии рибоизомеразы и ксилулозоэмпимеразы превращается в 2 молекулы рибозо-5-фосфата и 4 молекулы ксилулозо-5-фосфата
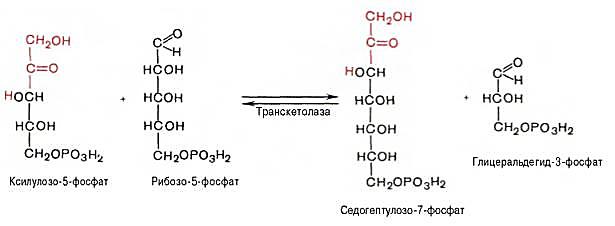
2 молекулы ксилулозо-5-фосфата взаимодействуют с 2 молекулами рибозо-5-фосфата в реакции, катализируемой транкетолазой. Коферментом транскетолазы является тиаминдифосфат. В результате реакций образуются 2 молекулы седогептулозо-7-фосфата, 2 молекулы 3-фосфоглицеральдегида и остается 2 молекулы ксилулозо-5-фосфата
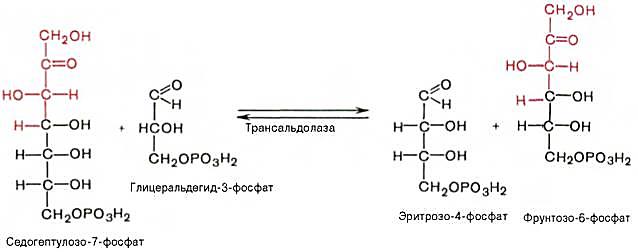
На следующем этапе происходит взаимодействие 2 молекул 3-фосфоглицеральдегида с 2 молекулами седогептулозо-7-фосфатом, что сопровождается образованием 2 молекул эритрозо-4-фосфата и фрутозо6-фосфата.
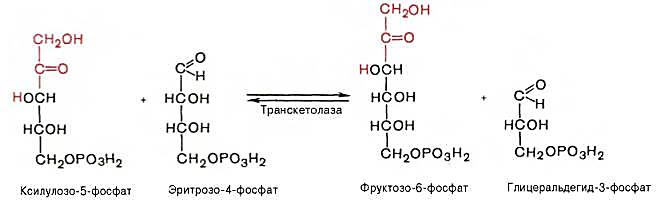
Оставшиеся 2 молекулы ксилулозо-5-фосфата вступают в транскетолазную реакцию с 2 молекулами эритрозо-4-фосфата, в результате образуются 2 молекулы фруктозо-6-фостфата и 2 молекулы 3-фосфоглицеральдегида
2 молекулы 3-фосфоглицеральдегида с участием альдолазы конденсируется в фруктозо-1,6,-дифосфат. Образовавшиеся 5 молекул фруктозо-6 фосфата изомеризуются в 5 молекул глюкозо-6-фосфата. С глюкозо-6-фосфата начинаются реакции пентозофосфатного пути и поэтому его можно отнести к циклическим путям метаболизма. Из 6 молекул глюкозо-6-фосфата, поступивших в пентозофосфатный цикл, 5 молекул снова регенерируют в это соединение, кроме того, образуются 6 молекул СО2 и 12 НАДФН(Н+).
|
|||||||
|
|
Последнее изменение этой страницы: 2021-05-27; просмотров: 113; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.108.9 (0.015 с.) |