Центральным в голографическом построении функций мозга является вопрос: на какой стадии системной организации поведенческого акта осуществляется интерференционное взаимодействие мотивационного и подкрепляющего возбуждений?
Проведенные нами эксперименты свидетельствуют о том, что голо- графические свойства мозга проявляются, прежде всего, на структурах акцептора результатов действия.
Доминирующая мотивация, будучи тесно связанной с акцептором результата действия, составляет с ним единую нейрональную архитектонику, в которой нейроны отражают как доминирующую потребность, так и свойства потребного будущего подкрепления.
Акцептор результата действия, тесно связанный с доминирующей мотивацией, широко представлен в структурах мозга. Он формируется за счет распространения копий исполнительных команд пирамидных нейронов коры мозга через коллатерали пирамидного тракта к вставочным нейронам, располагающимся на разных уровнях мозга: в коре и подкорковых структурах (Судаков К. В., 1984). К этим же нейронам по соответствующим афферентным каналам поступает множественная обратная афферентации от различных зрительных, обонятельных, вкусовых, тактильных и др. параметров подкрепляющих раздражителей - результатов, удовлетворяющих различные потребности организма (Рис. 7). На вставочных нейронах, образующих акцептор результата действия, и происходит взаимодействие мотивационных и подкрепляющих возбуждений (Sudakov K. V., 1998). Благодаря этому акцептор результата действия выступает в качестве своеобразного экрана, на котором разыгрываются голографические события взаимодействия доминирующей мотивации и подкрепления.
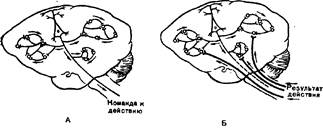
Рис. 7. Генерализованная локализация акцептора результата действия по структурам мозга. Объяснение в тексте. А - состояние до, Б - после получения информации от результата действия.
В ряде публикаций нами обоснована импринтинговая гипотеза формирования акцептора результата действия (Судаков К. В., 1964, 1987, 1996). Согласно этой гипотезе, подкрепляющие возбуждения всякий раз оставляют специфический след на ранее вовлеченных в архитектонику исходных доминирующих мотиваций структурах акцептора результата действия. Этот след и составляет энграмму подкрепления, своеобразную голограмму.
Специально проведенные нами опыты показали, что после предварительного подкрепления изменяются молекулярные свойства отдельных нейронов мозга, включенных в доминирующую мотивацию. Рибосомальный аппарат этих нейронов начинает синтезировать специальные белковые молекулы, определяющие их реакции на мотивационное возбуждение. Синтезируемые под влиянием доминирующей мотивации специфические белковые молекулы - эффекторные олигопептиды - участвуют в организации соответствующих поведенческих реакций. Эти процессы блокируются ингибиторами синтеза белка.
Подведение пентагастрина восстанавливает заблокированное циклогексимидом пищевое поведение и реакции нейронов сенсомоторной коры и дорзального гиппокампа на раздражение латерального гипоталамуса. Брадикинин восстанавливает оборонительную реакцию перепрыгивания кроликов через барьер, а АКТГ 4-10 - подавленную циклогексимидом реакцию самораздражения перифорникальной области гипоталамуса (Судаков С. К., 1987; Судаков К. В., 1992). Антитела (иммуноглобулины) к пентагастрину при их введении в боковые желудочки мозга также блокируют пищевые реакции у кроликов при раздражении латерального гипоталамуса (Судаков С. К., 1986).
Все это указывает на то, что в сформированных функциональных системах на основе предшествующих подкреплений строятся молекулярные энграммы поведения, на которые и оказывают действие блокаторы синтеза белка (Судаков К. В., 1995). Блокаторы синтеза белка нарушают сформированные предшествующими подкреплениями молекулярные энграммы, вследствие чего доминирующие мотивации при раздражении мотивациогенных структур мозга не приводят к синтезу эффекторных молекул и к результативному поведению животных.
Это, в свою очередь, позволяет думать, что синтезируемые на структурах акцептора результата действия под влиянием подкрепления и мотивации белковые молекулы участвуют в формировании голографического экрана мозга.
Молекулярная организация мозга при мотивации и подкреплении фиксируется определенными олигопептидами (Судаков К. В., 1992). Упорядоченные белковые взаимодействия, представленные по всему объему мозга, как показали наши опыты (Судаков К. В., 1987), могут сохраняться в течение более или менее длительного времени, составляя основу памяти.
Энграмма акцептора результата действия может быть разрушена введением антииммунных глобулинов к соответствующим белкам подкрепления. Ван В. Грейданус с соавт. (Van W. Greidanus et al., 1975) показали, что обучение у крыс нарушается после внутрижелудочковых инъекций антител к вазопрессину.
Можно полагать, что афферентная импульсация, поступающая в мозг от каждой потребности, когерентна микроколебаниям олигопептидов, экспрессируемых геномом составляющих акцептор результата действия нейронов мозга: молекулам пентагастрина при пищевой мотивации; молекулам брадикинина - при оборонительной мотивации и т.п. Вследствие этого мотивация, подобно опорной физической волне, на основе когерентных отношений и извлекает из памяти голограмму (энграмму) подкрепления.
Не исключено, что в объединении молекулярных энграмм подкрепления существенную роль играют также специфические мембранные белки, объединяемые адгезинами и захватывающие обширные участки мозговой ткани. Таким образом, энграммы подкрепления характеризуются новой молекулярной архитектоникой.
В специальных опытах (Анохин К.В. с соавт., 1988) у мышей линии CBLW мы исследовали роль блокатора синтеза ДНК, катализируемого обратной транскриптазой, - азидотимидина в формировании энграмм подкрепления. У мышей на основе обучения формировали навыки избегания темного отсека камеры при воздействии в нем электрическим током (Test step through). Полезным результатом мотивированных электрокожным раздражением животных в этом случае было нахождение животных в светлом отсеке камеры и предвидение наказующего действия темного отсека.
У другой группы мышей вырабатывали оборонительный навык нахождения на кубике (Test step down). При попытках спрыгнуть с кубиков животные получали электрокожное раздражение при контакте с полом клетки. Полезным результатом мотивированных электрокожным раздражением животных в этом случае было нахождение их на кубике и предвидение наказующего действия спрыгивания на пол. До и после сформирования указанных функциональных систем и соответствующих энграмм подкрепления животным вводили азидотимидин.
Введение азидотимидина в дозе 20 мкг/кг за час до начала обучения, а в других опытах в течение 3-х часов после завершения обучения выраженно нарушало у мотивированных страхом животных навык избегания темного отсека камеры. Несмотря на электрокожное раздражение, мыши начинали заходить в темный отсек. Эффект проявлялся до 7 суток. При введении азидотимидина через 5 часов после обучения указанного эффекта не наблюдалось. Аналогично азидотимидин блокировал выработку и сформированную функциональную систему нахождения животных на кубике - мыши начинали спрыгивать с кубика и получать электрокожное раздражение.
Приведенные опыты свидетельствуют о том, что блокатор синтеза ДНК - азидотимидин - препятствовал у животных выработке на основе электрокожных подкреплений энграммы подкрепления и, как следствие, - предвидения конечного приспособительного результата способов избегания повреждающих воздействий. Все это снова свидетельствует о том, что молекулярные энграммы памяти, сформированные на основе предшествующих подкреплений, тесно связаны с аппаратом акцептора результата действия.
Известно, что значительную часть генома нервных клеток составляют мозгоспецифические гистоновые и негистоновые белки хроматина.
В исследованиях (Мехтиев А. А. с соавт., 1985) кроликов в течение 6-8 месяцев иммунизировали негистоновыми белками хроматина, выделенными из мозга быков. Полученные в результате иммунизации кроликов антисыворотки и антитела вводили через вживленные канюли в боковые желудочки мозга экспериментальным крысам. Наибольшей активностью в регуляции мозгоспецифического синтеза РНК обладали антитела к ряду негистоновых белков хроматина Np-3,6, Np-8,6 (Np - нуклеопротеины с цифровым индексом, соответствующим их изоэлектрической точке).
Опыты показали, что введение крысам в боковые желудочки мозга антисыворотки к Np-3,6 и Np-8,6 через 24 часа после их обучения в челночной камере пассивному избеганию темного отсека, в котором они получали электрокожное раздражение, особенно при введении антисыворотки к Np-3,6 приводило к нарушению мотивированного страхом ранее выработанного у животных навыка пассивного избегания. Сокращалось число обученных животных, уменьшался латентный период захода крыс в темный отсек и общее время их пребывания в светлом отсеке камеры. Еще более выраженный эффект нарушения пассивной реакции избегания, особенно после введения антисыворотки к Np-8,6, наблюдали через 48 часов после предварительной выработки оборонительного навыка.
Проведенные опыты указывают на роль белков хроматина в опережающих механизмах энграммы подкрепления и предвидения животными повреждающих воздействий.
Все изложенное свидетельствует о том, что голографический экран акцептора результата действия на основе предшествующих подкреплений строится молекулярными механизмами генома нейрона. Кроме того, он обладает свойством опережающего отражения событий действительности. Можно думать, что в интеграции отдельных элементов акцептора результатов действия существенная роль принадлежит эмоции. Однако этот вопрос нуждается в специальном экспериментальном подтверждении.
Поскольку в организме постоянно взаимодействует множество функциональных систем в их иерархических, мультипараметрических и последовательных соотношениях, мозговая организация в целом представляет обобщенный голографический экран, на котором постоянно взаимодействует информация от внутренней и внешней среды. Однако, благодаря доминированию функциональных систем в каждом конкретном интервале времени в обобщенном голографическом экране доминирует акцептор результата деятельности ведущей функциональной системы, определяющей своей деятельностью ведущую потребность организма. Таким образом, мозговая организация динамически представлена сменяющими друг друга специфическими, голографическими экранами.





