Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Если гены а, в и с находятся в одной хромосоме, то генетические расстояния аддитивны (складываются и вычитаются), то есть расстояние ас равно либо сумме, либо разности расстояний ав и вс.⇐ ПредыдущаяСтр 17 из 17
Аддитивность расстояний является следствием линейного расположения генов в хромосомах. Если бы хромосомы ветвились, то АС совершенно не обязательно было бы суммой или разностью АВ и ВС: Таким образом, чисто гибридологически были установлены важные детали структуры генетического аппарата, задолго до открытия структуры ДНК и механизмов ее функционирования. Группы сцепления В эксперименте не может быть обнаружено сцепление между генами, расстояние между которыми превышает 50 сМ, так как максимальная доля кроссоверных потомков — 50% — наблюдается при полностью независимом наследовании. Однако хромосомы эукариот велики, содержат множество генов (каждая — тысячи генов), и многие из этих генов находятся на расстояниях, значительно превышающих 50% кроссинговера. Тем не менее, сцепление таких генов можно обнаружить косвенно, при посредстве генов, расположенных между ними. Например, в эксперименте установлено, что ген А сцеплен с В, В — с С, С— с D. Значит, ген А сцеплен и с D, хотя расстояние между ними столь велико, что в скрещивании сцепление заметить невозможно — гены ведут себя так, будто находятся в разных хромосомах и наследуются независимо. Это происходит потому, что расстояние между ними столь велико, что хотя бы 1 кроссинговер обязательно происходит на этом промежутке. Экспериментально было установлено, что число групп сцепления всегда совпадает с числом хромосом, что является одним из доказательств хромосомной теории наследственности.
4 группы сцепления и 4 хромосомы дрозофилы. Группы сцепления в ядерном геноме эукариот линейны, группы сцепления у прокариот и в геномах полуавтономных органелл — кольцевые. Поэтому особенности строения геномов разных организмов (линейная ДНК у эукариот, кольцевая — у прокариот) также были впервые установлены гибридологическими методами, без использования каких-либо молекулярных техник.
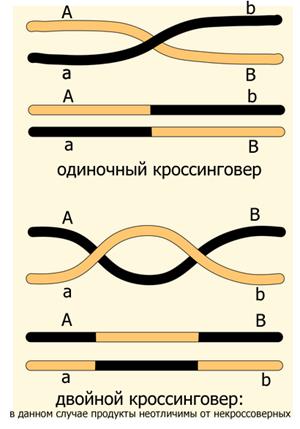
Карта кольцевой хромосомы кишечной палочки. Кратные кроссоверы При составлении генетических карт было замечено, что если гены А, В и С расположены в хромосоме в указанном порядке, то AC на самом деле никогда не равно в точности AВ+ВС, а несколько меньше, и этот эффект тем больше, чем больше расстояние между генами. Почему так происходит?
Дело в том, что если мы рассмотрим скрещивание цис-гетерозиготы AB/ab, то гаметы AB и ab мы считаем некроссоверными, а гаметы aB и Ab — кроссоверными, однако гаметы AB и ab могут образоваться не только без кроссинговера, но и в результате двойного (а также любого чётно-кратного — четверного и т.д.) кроссинговера, произошедшего между А и В. Поэтому экспериментально наблюдаемая частота кроссоверных потомков занижена — двойные кроссоверы в неё не входят.
Помочь делу может третий маркер — например, С — находящийся между А и В. Он позволяет заметить часть кратных кроссоверов и более точно установить расстояние между генами.
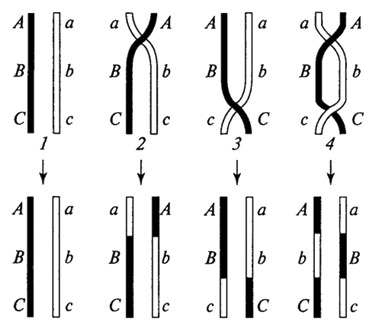
1 — без кроссинговера, 2 и 3 — одинарный кроссинговер, 4 — двойной кроссинговер Интерференция Влияние уже произошедшего акта кроссинговера на последующие кроссинговеры в близлежащих участках генома называется интерференцией. Она может быть как отрицательной (уменьшение вероятности нового кроссинговера по соседству с уже произошедшим), так и положительной. Как правило, наблюдается отрицательная интерференция, то есть подавление кроссинговера в близлежащих участках. Мерой интерференции является коэффициент совпадения С, или коэффициент коинциденции. Он равен отношению экспериментально наблюдаемой частоты кроссинговера к теоретически ожидаемой. а экспериментально наблюдаемая частота кроссинговера будет равна x = C*0.08% = 0.048% Строение нуклеиновых кислот В отличие от белков, углеводов и липидов, нуклеиновые кислоты никогда не накапливаются в клетке в больших количествах, и обнаружить их можно только с помощью специальных химических методов. Поэтому они были открыты только во второй половине XIX в., а по-настоящему изучить их роль в процессах жизнедеятельности удалось лишь во второй половине XX в. Так как первоначально они были обнаружены только в ядрах, им дали название нуклеиновые (от лат. nucleus — ядро).
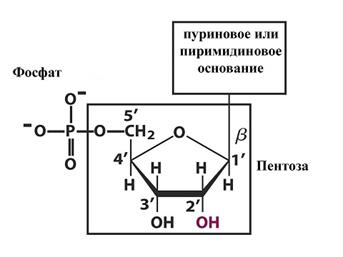
Нуклеиновые кислоты — биологические полимеры, мономерами которым служат нуклеотиды. Связи между нуклеотидами легко подвергаются гидролизу (распаду при реакции с водой). Каждый нуклеотид состоит из остатков углевода, фосфорной кислоты и азотистого основания (рис. 1).
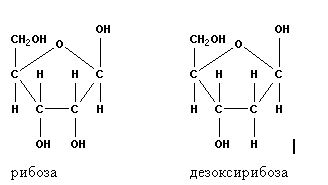
Рис. 1. Строение нуклеотида Углеводный компонент представлен пентозами — рибозой (в РНК) или дезоксирибозой (в ДНК), у которой отсутствует кислород при втором атоме углерода (рис. 2).
Рис. 2. Пентозы Остаток фосфорной кислоты образует сложноэфирную связь с гидроксилом при 5-м атоме углерода в сахаре. Соединение нуклеотидов в полимер происходит путем образования фосфатом одного нуклеотида второй эфирной связи с гидроксилом при 3-м углероде соседнего нуклеотида. Такая связь получила название фосфодиэфирной. Таким образом, нуклеиновые кислоты представляют собой цепь из чередующихся остатков пентозы и фосфорной кислоты (рис. 3). Кроме того, от первого атома углерода каждой пентозы отходит в бок азотистое основание. В этом нуклеиновые кислоты сходны с белками, в которых полимерная цепь образована пептидными группировками с отходящими от них боковыми радикалами аминокислот. Так же, как и у белков, в нуклеиновых кислотах два конца цепи неодинаковы. С одной стороны имеется не занятое связью пятое положение рибозы, этот конец называют 5’-концом. С противоположной стороны не занят связью третий гидроксил сахара, этот конец обозначают как 3’-конец. 5’-конец считается началом цепи, а 3’-конец — ее окончанием.
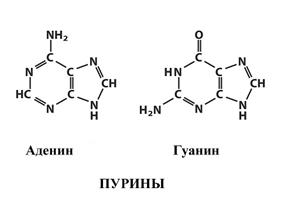
Рис. 3. Нуклеиновые кислоты В одной молекуле нуклеиновой кислоты присутствует только один вид пентозы. Те молекулы, которые содержат рибозу, называют рибонуклеиновой кислотой, или сокращенно РНК. Нуклеиновую кислоту, содержащие дезоксирибозу, называют дезоксирибонуклеиновой кислотой, или ДНК. Помимо пентозы, нуклеиновые кислоты отличаются азотистыми основаниями. Они представляют собой ароматические циклы, содержащие несколько атомов азота и заместители при определенных атомах углерода. По структуре гетероциклов азотистые основания делятся на две группы. Пиримидиновые азотистые основания: урацил, тимин и цитозин (рис. 4). Тимин отличается от урацила только наличием метильной группы, что незначительно меняет его свойства. В РНК встречаются урацил и цитозин, а в ДНК — тимин и цитозин. Пуриновые основания: аденин и гуанин (рис. 5). Во всех нуклеиновых кислотах присутствуют оба пурина.
Рис. 4. Пиримидиновые основания Рис. 5. Пуриновые основания За счет чередования различных нуклеотидов в цепи нуклеиновые кислоты могут достигать огромного многообразия (количество видов полимеров равно числу видов мономеров в степени, равной числу мономеров в цепи). И хотя число мономеров в нуклеиновых кислотах меньше, чем в белках, степень полимерности, особенно у ДНК, намного выше. Длина цепей ДНК, входящих в хромосомы разных организмов, составляет от миллионов до сотен миллионов нуклеотидов.
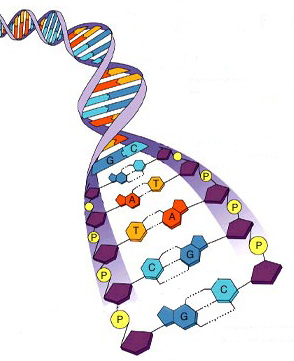
Молекулы РНК обычно короче, их длина — от нескольких десятков до нескольких десятков тысяч нуклеотидов. А при длине цепи 500 нуклеотидов количество возможных комбинаций составляет более 10 300. Принцип комплЕментарности При анализе содержания азотистых оснований в ДНК из различных организмов Эрвин Чаргафф обнаружил определенные закономерности, позднее названные правилами Чаргаффа. Молярное содержание аденина всегда равно молярному содержанию тимина, а молярное содержание гуанина — молярному содержанию цитозина. Количество пуринов равнялось количеству пиримидинов, а отношение А+Т/Г+Ц было различным у разных видов живых организмов. Это указывало на возможные взаимодействия оснований в ДНК между собой. На основании правил Чаргаффа и предварительных результатов рентгеноструктурного анализа Джеймс Уотсон и Френсис Крик в 1953 г. предложили двуспиральную модель структуры ДНК. Согласно этой модели молекула ДНК состоит из двух полинуклеотидных цепей, соединенных между собой азотистыми основаниями. При этом аденин одной цепи всегда взаимодействует с тимином в другой, и наоборот. Точно так же гуанин одной цепи всегда связан с цитозином в другой (рис. 6).
Рис. 6. Образование водородных связей между азотистыми основаниями
Главной особенностью пар А–Т и Г–Ц является их одинаковая геометрия. Это позволяет построить двуспиральную молекулу с постоянным расстоянием между цепями, построенными остатками сахара и фосфорной кислоты. Образование любых других пар приводит к нарушению правильной структуры. Такое взаимодействие оснований, при котором они дополняют друг друга до определенной структуры, одинаковой для всех пар, получило название принципа комплементарности. Пары аденин и тимин, гуанин и цитозин называются комплементарными парами, а две цепочки нуклеиновых кислот, в которых все основания образуют комплементарные пары — комплементарными цепочками. Таким образом, каждая молекула ДНК состоит из двух комплементарных цепочек полинуклеотидов (рис. 7).
Рис. 7. Принцип комплиментарности Важной особенностью структуры двойной спирали ДНК является то, что комплементарные цепи направлены в противоположные стороны, т. е. 5’-конец одной цепи связан комплементарными основаниями с 3’-концом другой цепи, и наоборот. Основания плотно слипаются своими плоскостями, что делает связь между цепочками еще более прочной. Такое слипание получило название стэкинг-взаимодействия. В результате в центре молекулы ДНК находится как бы стержень, построенный из азотистых оснований, а по краям он обвит двумя нитями, состоящими из чередующихся остатков дезоксирибозы и фосфорной кислоты.
Сравнение ДНК и РНК
Рис. 8. Различия в строении ДНК и РНК
|
||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-02-07; просмотров: 127; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.64.235 (0.022 с.) |