Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Локализация медиаторов и соответствующих нейронов ЦНС.
В разных отделах ЦНС расположены нейроны, имеющие различные медиаторы. Ацетилхолин является медиатором альфа-мотонейронов спинного мозга и ствола (их аксоны заканчиваются нервно-мышечным синапсом), нейронов коры больших полушарий, ретикулярной формации мозга, преганглионарных нейронов вегетативной (симпатической и парасимпатической) нервной системы, ганглионарных нейронов парасимпатической нервной системы, а также нейронов сетчатки. Дофамин является медиатором нейронов, сконцентрированных в среднем мозге (черная субстанция, покрышка мозга), в гипоталамусе, в симпатических ганглиях и в сетчатке. Аксоны дофаминергических нейронов достигают нейронов базальных ганглиев, лимбической системы, коры больших полушарий. Норадреналин является медиатором нейронов, локализованных в голубом пятне ствола мозга (с проекцией в кору мозга, гипоталамус, мозжечок, спинной мозг), а также ганглионарных нейронов симпатической нервной системы. Серотонин является медиатором серотонинергических нейронов, локализованных, главным образом, в ядрах шва ствола мозга (проекция аксонов в кору головного мозга, гипоталамус, мозжечок, спинной мозг) и в сетчатке. Гистамин является медиатором нейронов сосцевидных телец гипоталамуса. Аксоны гистаминергических нейронов проецируются в кору мозга, таламус, базальные ганглии, мозжечок, спинной мозг. Глютамат является медиатором возбуждающих нейронов, локализованных в различных отделах коры больших полушарий, в таламусе, базальных ганглиях, мозжечке, гипоталамусе, стволе мозга, спинном мозге, а также в сетчатке. Глицин является медиатором тормозных нейронов, расположенных главным образом в спинном мозге и в сетчатке. ГАМК является медиатором тормозных нейронов коры мозга, мозжечка, ствола мозга, спинного мозга (совместно с глицином) и сетчатки. АТФ является медиатором возбуждающих нейронов уздечки головного мозга, спинного мозга, афферентных нейронов, а также симпатических нейронов. В роли медиаторов могут выступать и нейропептиды. Для них характерна следующая локализация в ЦНС. Субстанция Р - головной мозг, окончания афферентных нейронов в ноцицептивной (отвечающей за передачу болевых ощущений) системе. Вазопрессин, окситоцин - задний гипофиз, продолговатый мозг, спинной мозг. Кортиколиберин - медиальное возвышение гипоталамуса и другие отделы мозга. Тиреолиберин - гипоталамус, сетчатка. Соматостатин - гипоталамус и другие отделы мозга, желатинозная субстанция, сетчатка. Гонадолиберин - гипоталамус, хеморецептивные зоны желудочков мозга, преганглионарные окончания, сетчатка. Эндотелин - задний гипофиз, ствол мозга. Энкефалины - желатинозная субстанция, многие другие отделы ЦНС, сетчатка. Эндорфины - гипоталамус, ствол мозга, сетчатка. Вазоактивный интестинальный полипептид - ганглионарные холинергические нейроны, некоторые чувствительный нейроны, гипоталамус, кора больших полушарий головного мозга, сетчатка. Нейротензин - гипоталамус, сетчатка. Гастрин - гипоталамус, продолговатый мозг. Глюкагон - гипоталамус, сетчатка. Мотилин - гипоталамус, таламус, обонятельная луковица, ствол мозга, перегородка, гиппокамп, стриатум. Пептид, генетически родственный кальцитонину - окончания первичных афферентов, вкусовой анализатор. Нейропептид Y - симпатические нейроны вегетативной нервной системы. Ангиотензин II – гипоталамус, миндалевидное тело, ствол мозга. Галанин - гипоталамус. Атриопептин - гипоталамус, ствол мозга.
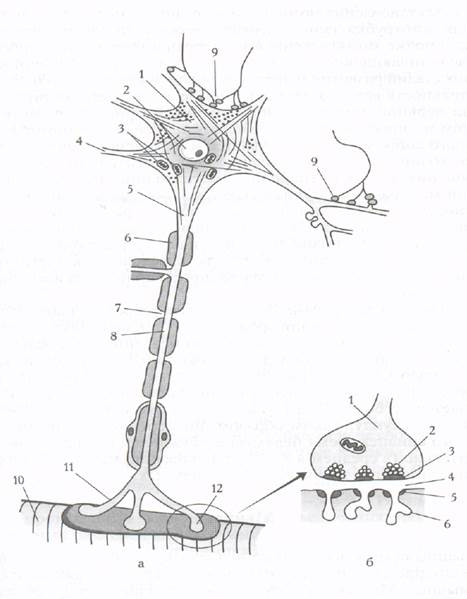
Нервно-мышечный синапс Структурно-функциональное созревание нервно-мышечных синапсов охватывает почти весь период антенатального и раннего постнатального периодов развития. Оно имеет свои особенности для пресинапса и постсинапса (пре- и постсинаптических мембран). Строение нервно-мышечного синапса представлено на рис.4. Рассмотрим созревание пресинапса. Двигательные нервные окончания в мышцах появляются на 13- 14-й неделе внутриутробного развития. Формирование их продолжается длительное время и после рождения. Мышечные волокна новорожденного, как и взрослого человека, «монотерминальны», т.е. имеют один синапс в виде типичной концевой бляшки. Созревание пресинапса проявляется увеличением терминального разветвления аксона, усложнением его формы, увеличением площади всего окончания. Степень созревания нервных окончаний значительно увеличивается к 7-8 годам; при этом появляются более быстрые и разнообразные движения. Однако и в данном возрасте еще не достигается степень развития движений, наблюдаемая у взрослых.
Мышцы с преимущественно динамическими функциями, обеспечивающие точные разнообразные движения, имеют более многообразные и сложные по форме концевые разветвления в двигательных бляшках, чем мышцы, обеспечивающие однообразные движения. В процессе развития усиливается синтез ацетилхолина в мотонейронах, возрастает количество активных зон в пресинаптическом окончании и количество квантов медиатора, выделяющегося в синаптическую щель. Созревание постсинапса: когда окончание аксона достигает миотрубки, в соответствующих ее участках появляется скопление митохондрий, рибосом, трубчатых образований. На поверхности миотрубки образуются выемки, в которых помещается окончаний аксона. Формируется примитивное нервно-мышечное соединение.
Рис.4. Строение нейрона с нервно-мышечным синапсом (схема): а - строение нейрона: 1 - тело нейрона, 2 - ядро, 3 - митохондрии, 4 - дендрит, 5 - аксон, 6 - миелиновая оболочка, 7 - перехват Ранвье, 8 - аксоплазма, 9 - синаптическое окончание, 10 - мышца, 11 - нервное окончание, 12 - нервно-мышечный синапс; б - строение нервно-мышечного синапса: 1 - пресинаптическое окончание, 2 - синаптические пузырьки, содержащие ацетилхолин, 3 - пресинаптическая мембрана, 4 - синаптическая щель, 5 - холинорецепторы на постсинап- тической мембране, 6 - складки на постсинаптической мембране
В ходе становления моторной иннервации существует этап, когда каждая миотрубка (мион) имеет множественную иннервацию. Однако позже множественная иннервация устраняется в процессе увеличивающейся двигательной активности. Особенностью ранних стадий развития мышечных волокон является разлитая чувствительность всей поверхности мембраны к ацетилхолину, присущая нервной стадии развития мышц. В процессе развития на каждом мионе сохраняется единственный синапс, формирование которого сопровождается появлением в постсинаптической мембране холинэстеразы. Увеличение ее концентрации приводит к повышению скорости гидролиза ацетилхолина. На постсинаптической мембране образуются складки, происходит концентрация холинорецепторов в синаптической зоне, в результате чего увеличивается амплитуда потенциала концевой пластинки (ПКП) и повышается надежность передачи возбуждения через синапс. Внесинаптическая поверхность мышечного волокна постепенно теряет чувствительность к ацетилхолину. После денервации она вновь распространяется на всю поверхность мембраны. Вследствие незрелости нервно-мышечного синапса у плода и новорожденного синаптическая передача возбуждения происходит относительно медленно: у взрослых длительность синаптической задержки около 0,5 мс, у новорожденных - около 4,5 мс. Соответственно низка и лабильность синапсов новорожденных. Без трансформации ритма через такой синапс передается не более 20 импульсов, а у взрослых – 100-150 импульсов в секунду. Таким образом, по мере созревания мионеврального синапса время перехода возбуждения с нерва на мышцу значительно (в среднем в 8 -10 раз) укорачивается и к 7 -8 годам жизни становится таким же, как у взрослых.
|
||||||
|
|
Последнее изменение этой страницы: 2019-11-02; просмотров: 166; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.143.4.181 (0.007 с.) |