Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Некоторые особенности функционирования нервной системы человекаСтр 1 из 17Следующая ⇒
Н.В.Мамылина Некоторые особенности функционирования нервной системы человека Челябинск 2007
В данном учебно-методическом пособии рассмотрены анатомические, физиологические и возрастные особенности нервной системы человека: строение нервной ткани, нервных волокон, нейронов; проведение возбуждения по нервным волокнам; свойства нервных центров; строение и принципы функционирования синапсов. Дано общее представление о рефлекторной дуге, рефлекторном кольце и принципах координационной деятельности ЦНС. В данном учебно-методическом пособии акцентируется внимание на возрастные особенности структуры и функции нервных клеток. Данное учебно-методическое пособие подготовлено в соответствии с Государственным образовательным стандартом Высшего образования и предназначено для студентов факультета физической культуры и БЖ, а также РИПОДО. Рецензент: доктор биологических наук, профессор ЮУрГУ Попова Т.В. Введение Анатомия и физиология- это важнейшие науки о строении и функциях человеческого организма. Знать «устройство», отличительные особенности органов, систем, тканей, клеток организма человека должен каждый педагог по физической культуре, безопасности жизнедеятельности, дошкольной педагогике, логопедии и др. Человек как представитель животного мира, подчиняется биологическим закономерностям, присущим всем живым существам. Но отличительной особенностью человека является высшая нервная деятельность, высокоразвитый интеллект, наличие членораздельной речи, социальные условия жизни и многое другое. Знание особенностей строения и функций человеческого организма полезно любому человеку, так как может возникнуть потребность оказать первую медицинскую помощь (остановить кровотечение, сделать искусственное дыхание и т.п.). Знание анатомии и физиологии с возрастными особенностями организма даёт возможность разрабатывать гигиенические нормы, необходимые в быту в целях сохранения здоровья человека. В данном учебно-методическом пособии уделяется внимание возрастным особенностям систем организма человека. Это даст возможность понять механизмы происходящих в организме в различные возрастные периоды процессов, изучить взаимосвязь человека с внешней средой, аномалий, пороков развития.
Данное пособие является одним из серии пособий по возрастным и анатомо-физиологическим особенностям органов и систем организма человека. Пособие содержит новые научные знания и факты, выявляющие основные закономерности развития нервной системы организма человека. Совершенствование методов исследования, широкое использование электронного микроскопа, научные достижения в области молекулярной биологии, биофизики, биохимии, генетики позволили обобщить имеющиеся в пособии научные факты.
Классификация нейронов Существует большое многообразие нейронов ЦНС. Поэтому предложены и различные варианты их классификации. Чаще всего эта классификация осуществляется по трем признакам - морфологическим, функциональным и биохимическим. Морфологическая классификация нейронов учитывает количество отростков у нейронов и подразделяет все нейроны на три типа - униполярные, биполярные и мультиполярные. Униполярные нейроны (от лат. унус - один; синонимы - одноотростчатые, или однополюсные, нейроны) имеют один отросток. По мнению одних исследователей, в нервной системе человека и других млекопитающих нейроны этого типа не встречаются. Однако некоторые авторы полагают, что униполярные нейроны отмечаются у человека в период раннего эмбрионального развития, а в постнатальном онтогенезе они встречаются в мезэнцефалическом ядре тройничного нерва (обеспечивают проприоцептивную чувствительность жевательных мышц). Ряд исследователей к униполярным клеткам относят амакриновые нейроны сетчатки глаза и межклубочковые нейроны обонятельной луковицы. Биполярные нейроны (синонимы - двухотросчатые, или двухполюсные, нейроны) имеют два отростка - аксон и дендрит, обычно отходящие от противоположных полюсов клетки. В нервной системе человека собственно биполярные нейроны встречаются в основном в периферических частях зрительной, слуховой и обонятельной систем, например, биполярные клетки сетчатки глаза, спирального и вестибулярного ганглиев. Биполярные нейроны дендритом связаны с рецептором, аксоном - с нейроном следующего уровня организации соответствующей сенсорной системы.
Однако значительно чаще в ЦНС человека и других животных встречается разновидность биполярных нейронов - так называемые псевдоуниполярные, или ложноуниполярные, нейроны. У них оба клеточных отростка (аксон и дендрит) отходят от тела клетки в виде единого выроста, который далее Т-образно делится на дендрит и аксон: первый идет с периферии от рецепторов, второй направляется в ЦНС. Эти клетки встречаются в сенсорных спинальных и краниальных ганглиях. Они обеспечивают восприятие болевой, температурной, тактильной, проприоцептивной, барорецептивной и вибрационной сигнализации. Мультиполярные нейроны имеют один аксон и много (2 и более) дендритов. Они наиболее распространены в нервной системе человека. Описано до 60-80 вариантов этих клеток. Однако все они представляют разновидности веретенообразных, звездчатых, корзинчатых, грушевидных и пирамидных клеток. По длине аксона выделяют клетки Гольджи I типа (с длинным аксоном) и клетки Гольджи II типа (с коротким аксоном). С точки зрения локализации нейронов их можно разделить на нейроны ЦНС, т.е. находящиеся в спинном (спинальные нейроны) и головном мозге (бульбарные, мезенцефальные, церебеллярные, гипоталамические, тала-мические, корковые), а также за пределами ЦНС, т.е. входящие в состав периферической нервной системы - это нейроны вегетативных ганглиев, а также нейроны, составляющие основу метасимпатического отдела вегетативной нервной системы. Функциональная классификация нейронов разделяет их по характеру выполняемой ими функции (в соответствии с их местом в рефлекторной дуге) на три типа: афферентные (чувствительные), эфферентные (двигательные) и ассоциативные. 1. Афферентные нейроны (синонимы - чувствительные, рецепторные, центростремительные), как правило, являются ложноуниполярными нервными клетками. Тела этих нейронов располагаются не в ЦНС, а в спинномозговых узлах или чувствительных узлах черепно-мозговых нервов. Один из отростков, отходящий от тела нервной клетки, следует на периферию, к тому пли иному органу и заканчивается там сенсорным рецептором, который способен трансформировать энергию внешнего стимула (раздражения) в нервный импульс. Второй отросток направляется в ЦНС (спинной мозг) в составе задних корешков спинномозговых нервов или соответствующих чувствительных волокон черепно-мозговых нервов. Как правило, афферентные нейроны имеют небольшие размеры и хорошо разветвленный на периферии дендрит. Функции афферентных нейронов тесно связаны с функциями сенсорных рецепторов. Таким образом, афферентные нейроны генерируют нервные импульсы под влиянием изменений внешней или внутренней среды Часть нейронов, принимающих участие в обработке сенсорной информации, которые можно рассматривать как афферентные нейроны высших отделов мозга, принято делить в зависимости от чувствительности к действию раздражителей на моносенсорные, бисенсорные и полисенсорные. Моносенсорные нейроны располагаются чаще в первичных проекционных зонах коры и реагируют только на сигналы своей сенсорности. Например, значительная часть нейронов первичной зоны зрительной области коры полушарий головного мозга реагирует только на световое раздражение сетчатки глаза.
Моносенсорные нейроны подразделяют функционально по их чувствительности к разным качествам одного раздражителя. Так, отдельные нейроны слуховой зоны коры больших полушарий головного мозга могут реагировать на предъявления тона 1000 Гц и не реагировать на тоны другой частоты. Они называются мономодальными. Нейроны, реагирующие на два разных тона, называются бимодальными, на три и более - полимодальными. Бисенсорные нейроны чаще располагаются во вторичных зонах коры какого-либо анализатора и могут реагировать на сигналы как своей, так и другой сенсорности. Например, нейроны вторичной зоны зрительной области коры больших полушарий головного мозга реагируют на зрительные и слуховые раздражения. Полисенсорные нейроны - это чаще всего нейроны ассоциативных зон мозга; они способны реагировать на раздражение слуховой, зрительной, кожной и других рецептивных систем. 2. Эфферентные нейроны (синонимы - двигательные, моторные, секреторные, центробежные, сердечные, сосудодвигательные и пр.) предназначены для передачи информации от ЦНС на периферию, к рабочим органам. Например, эфферентные нейроны двигательной зоны коры большого мозга - пирамидные клетки - посылают импульсы к альфа-мотонейронам передних рогов спинного мозга, т.е. они являются эфферентными для этого отдела коры большого мозга. В свою очередь альфа-мотонейроны спинного мозга являются эфферентными для его передних рогов и посылают сигналы к мышцам. По своему строению эфферентные нейроны - это мультиполярные нейроны, тела которых находятся в сером веществе ЦНС (или на периферии в вегетативных узлах различных порядков). Аксоны этих нейронов продолжаются в виде соматических или вегетативных нервных волокон (периферических нервов) к соответствующим рабочим органам, в том числе к скелетным и гладким мышцам, а также к многочисленным железам. Основной особенностью эфферентных нейронов является наличие длинного аксона, обладающего большой скоростью проведения возбуждения. Эфферентные нейроны разных отделов коры больших полушарий связывают между собой эти отделы по аркуатным связям. Такие связи обеспечивают внутриполушарные и межполушарные отношения. Все нисходящие пути спинного мозга (пирамидный, руброспинальный, ретикулоспинальный и т.д.) образованы аксонами эфферентных нейронов соответствующих отделов ЦНС. Нейроны автономной нервной системы, например, ядер блуждающего нерва, боковых рогов спинного мозга также относятся к эфферентным нейронам.
3. Вставочные нейроны (синонимы - интернейроны, контактные, ассоциативные, коммуникативные, объединяющие, замыкательные, проводниковые, кондукторные) осуществляют передачу нервного импульса с афферентного (чувствительного) нейрона на эфферентный (двигательный) нейрон. Суть этого процесса состоит в передаче полученного афферентным нейроном сигнала эфферентному нейрону для исполнения в виде ответной реакции организма. И. П. Павлов определил сущность этого как «явление нервного замыкания». Вставочные нейроны располагаются в пределах серого вещества ЦНС. По своему строению - это мультиполярные нейроны. Считается, что в функциональном отношении это наиболее важные нейроны ЦНС, так как на их долю приходится 97 %, а по некоторым данным, - даже 98-99 % от общего числа нейронов ЦНС. Область влияния вставочных нейронов определяется их строением, в том числе длиной аксона и числом коллатералей. Например, многие вставочные нейроны имеют аксоны, которые заканчиваются на нейронах своего же центра, обеспечивая, прежде всего, их интеграцию. Одни вставочные нейроны получают активацию от нейронов других центров и затем распространяют эту информацию на нейроны своего центра. Это обеспечивает усиление влияния сигнала за счет его повторения в параллельных путях и удлиняет время сохранения информации в центре. В итоге центр, куда пришел сигнал, повышает надежность воздействия на исполнительную структуру. Другие вставочные нейроны получают активацию от коллатералей эфферентных нейронов своего же центра и затем передают эту информацию назад в свой же центр, образуя обратные связи. Так организуются реверберирующие сети, позволяющие длительно сохранять информацию в нервном центре. Вставочные нейроны по своей функции могут быть возбуждающими или тормозными. При этом возбуждающие нейроны могут не только передавать информацию с одного нейрона на другой, но и модифицировать передачу возбуждения, в частности, усиливать ее эффективность. Например, в коре большого мозга имеются «медленные» пирамидные нейроны, которые влияют на активность «быстрых» пирамидных нейронов. Очевидно, что среди вставочных нейронов можно также выделить командные нейроны, песймекерные, гормонпродуцирующие нейроны (например, нейроны тубероинфундибулярной области гипоталамуса), потребностно-мотивационные, гностические и многие другие виды нейронов. Биохимическая классификация нейронов основана на химических особенностях нейромедиаторов, используемых нейронами в синаптической передаче нервных импульсов. Выделяют много различных групп нейронов, в частности, холинергические (медиатор - ацетилхолин), адренергические (медиатор - норадреналин), серотонинергические (медиатор - серотонин), дофаминергические (медиатор - дофамин), ГАМК-ергические (медиатор - гамма-аминомасляная кислота - ГАМК), пуринергические (медиатор - АТФ и его производные), пептидергические (медиаторы - субстанция Р, энкефалины, эндорфины, вазоактивный интестинальный пептид, холецистокинин, нейротензин, бомбезин и другие нейропептиды). В некоторых нейронах терминали содержат одновременно два типа нейромедиатора, а также нейромодуляторы.
Распределение нейронов, использующих различные медиаторы, в нервной системе неравномерно. Нарушение выработки некоторых медиаторов в отдельных структурах мозга связывают с патогенезом ряда нервно-психических заболеваний. Так, содержание дофамина снижено при паркинсонизме и повышено при шизофрении, снижение уровня норадреналина и серотонина типично для депрессивных состояний, а их повышение - для маниакальных. Нейроны, продуцирующие гормоны, можно также разделить по группам, в зависимости от природы продуцируемого ими гормона (кортиколиберин-, гонадолиберин-, тиролиберинпродуцирующие, пролактостатинпродуцирующие и другие). Другие виды классификаций нейронов. Нервные клетки разных отделов нервной системы могут быть активными вне воздействия, т.е. обладают свойством автоматии. Их называют фоновоактивными нейронами. Другие нейроны проявляют импульсную активность только в ответ на какое-либо раздражение, т.е. они не обладают фоновой активностью. Некоторые нейроны, по причине их особой значимости в деятельности мозга, получили дополнительные названия по имени исследователя, впервые описавшего соответствующие нейроны. Среди них - пирамидные клетки Беца, локализованные в новой коре большого мозга; грушевидные клетки Пуркинье, клетки Гольджи, клетки Лугано (все - в составе коры мозжечка); тормозные клетки Реншоу (спинной мозг) и ряд других нейронов.
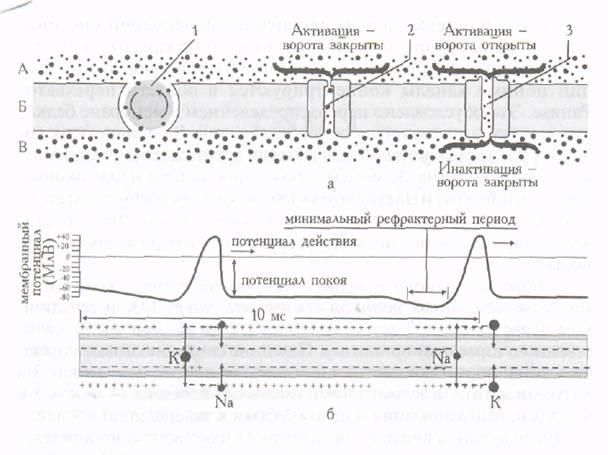
Свойства нервных волокон Мембрана аксона разделяет жидкие среды, различающиеся по содержанию ионов натрия и калия (в жидкой среде, окружающей аксон, в 10 раз больше ионов натрия, чем ионов калия, внутри аксона - обратное соотношение). Встроенные в аксональную мембрану протеины играют роль селективных ионных каналов, пропускающих через мембрану ионы натрия или калия. В состоянии покоя, когда передача нервного импульса отсутствует, оба вида каналов закрыты и ионный (натрий-калиевый насос) поддерживает неравномерное (указанное выше) соотношение ионов натрия и калия, выкачивая наружу ионы натрия в обмен на ионы калия. Внутренняя поверхность мембраны в состоянии покоя заряжена отрицательно (приблизительно на 70 мВ) по отношению наружной мембране. При уменьшении заряда вследствие возникновения нервного импульса открываются натриевые каналы, пропускающие ионы натрия внутрь аксона. Вслед за этим натриевые каналы закрываются и открываются калиевые, выпускающие на наружную поверхность мембраны ионы калия. Ионные каналы последовательно открываются и закрываются, что обеспечивает распространение нервного импульса (рис.2).
Рис.2. Строение мембраны аксона (а) и распространение нервного импульса по аксону (б). Ионы натрия обозначены мелкими точками, ионы калия - крупными точками. А - наружная сторона мембраны аксона, Б - мембрана аксона, В - внутренняя сторона мембраны аксона. 1 - ионный насос, 2 - калиевый канал, 3 - натриевый канал
Распространению нервного импульса вдоль аксона способствует локализованное поступление внутрь аксона ионов натрия с последующим выходом ионов калия через каналы, которые открываются и закрываются под влиянием изменения разности потенциалов. Зарождение нервного импульса, который далее распространяется по аксону, происходит в теле нейрона. При небольшом изменении разности потенциалов открываются отдельные натриевые каналы, что еще больше увеличивает разность потенциалов. В месте поступления ионов натрия внутрь аксона внутренняя мембрана становится положительно заряженной. Возникающие изменения разности потенциалов приводят к тому, что натриевые каналы закрываются, открываются калиевые, калий выходит на наружную поверхность мембраны. Результатом этого процесса является быстрое восстановление отрицательного заряда внутренней мембраны. Изменение разности потенциалов носит название «потенциал действия». Потенциал действия распространяется по аксону. После короткого рефрактерного периода может возникать второй импульс. Возбудимость нервных волокон плода и новорожденного значительно ниже, чем у взрослого, но с 3-месячного возраста она начинает повышаться. Величина хронаксии в несколько раз больше, чем у взрослых. Потенциал покоя у детей значительно меньше, чем у взрослых, из-за большой проницаемости клеточной мембраны для ионов. В процессе созревания нервного волокна проницаемость его мембраны уменьшается, что ведет к возрастанию ионной асимметрии. Последняя и является непосредственной причиной увеличения потенциала покоя. При этом увеличивается и потенциал действия, что также свидетельствует о функциональной зрелости нервного волокна. Небольшая величина потенциала действия новорожденного сочетается с большой его продолжительностью и часто с отсутствием реверсии, что объясняется меньшей, чем у взрослого, ионной асимметрией. Фазовые изменения возбудимости во время возбуждения в раннем постнатальном онтогенезе также имеют особенности. В частности, длительность абсолютной рефлекторной фазы 5-8 мс, а относительной – 40-60 мс (у взрослых - 0,5-2 и 2 -10 мс соответственно). Проводимость нерва плода и детей низкая. У новорожденных, например, скорость проведения возбуждения по нервным волокнам не превышает 50 % скорости взрослых. Так, в локтевом нерве максимальная скорость составляет у взрослых 62 м/с, у новорожденных - 31 м/с. Плохо выражена изолированность проведения возбуждения. С возрастом скорость проведения возбуждения по нервным волокнам возрастает в результате их миелинизации, увеличения толщины нервного волокна и его потенциала действия. Миелинизация нервных волокон обеспечивает увеличение скорости проведения возбуждения вследствие того, что непрерывное проведение возбуждения сменяется сальтаторным и увеличивается расстояние между перехватами Ранвье (потенциал действия «перепрыгивает» на большее расстояние). Увеличение толщины безмякотного нервного волокна и еще не покрытого миелином мякотного волокна ведет к ускорению проведения возбуждения в результате уменьшения продольного сопротивления ионному току в аксоплазме. При увеличении же мембранного потенциала проведение возбуждения ускоряется, потому что большой потенциал действия быстрее вызывает возбуждение соседнего участка нервного волокна. Скорость распространения возбуждения по нервным волокнам у детей становится такой же, как у взрослых, к 5-9 годам, чтo связано с завершением миелинизации различных волокон в разные сроки и окончанием увеличения диаметров осевых цилиндров. Лабильность - функциональная подвижность нервной ткани (термин впервые сформулировал Н.Е.Введенский)- определяется максимальным ритмом возбуждения, который ткань может воспроизводить в соответствии с ритмом стимулов. Лабильность свидетельствует о скорости метаболических процессов в нервной ткани и обусловливает характер возбуждения и торможения в ней, лабильность нервного волокна у плода и детей первых лет жизни низкая. С возрастом она увеличивается: число потенциалов действия, которое способно воспроизвести волокно в 1 с у новорожденных, например, составляет 4-10, а у детей 5-9 лет приближается к норме взрослых (300-1000 импульсов).
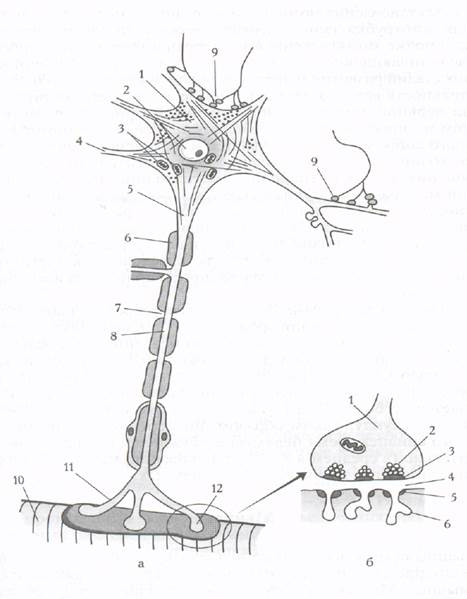
Нервно-мышечный синапс Структурно-функциональное созревание нервно-мышечных синапсов охватывает почти весь период антенатального и раннего постнатального периодов развития. Оно имеет свои особенности для пресинапса и постсинапса (пре- и постсинаптических мембран). Строение нервно-мышечного синапса представлено на рис.4. Рассмотрим созревание пресинапса. Двигательные нервные окончания в мышцах появляются на 13- 14-й неделе внутриутробного развития. Формирование их продолжается длительное время и после рождения. Мышечные волокна новорожденного, как и взрослого человека, «монотерминальны», т.е. имеют один синапс в виде типичной концевой бляшки. Созревание пресинапса проявляется увеличением терминального разветвления аксона, усложнением его формы, увеличением площади всего окончания. Степень созревания нервных окончаний значительно увеличивается к 7-8 годам; при этом появляются более быстрые и разнообразные движения. Однако и в данном возрасте еще не достигается степень развития движений, наблюдаемая у взрослых. Мышцы с преимущественно динамическими функциями, обеспечивающие точные разнообразные движения, имеют более многообразные и сложные по форме концевые разветвления в двигательных бляшках, чем мышцы, обеспечивающие однообразные движения. В процессе развития усиливается синтез ацетилхолина в мотонейронах, возрастает количество активных зон в пресинаптическом окончании и количество квантов медиатора, выделяющегося в синаптическую щель. Созревание постсинапса: когда окончание аксона достигает миотрубки, в соответствующих ее участках появляется скопление митохондрий, рибосом, трубчатых образований. На поверхности миотрубки образуются выемки, в которых помещается окончаний аксона. Формируется примитивное нервно-мышечное соединение.
Рис.4. Строение нейрона с нервно-мышечным синапсом (схема): а - строение нейрона: 1 - тело нейрона, 2 - ядро, 3 - митохондрии, 4 - дендрит, 5 - аксон, 6 - миелиновая оболочка, 7 - перехват Ранвье, 8 - аксоплазма, 9 - синаптическое окончание, 10 - мышца, 11 - нервное окончание, 12 - нервно-мышечный синапс; б - строение нервно-мышечного синапса: 1 - пресинаптическое окончание, 2 - синаптические пузырьки, содержащие ацетилхолин, 3 - пресинаптическая мембрана, 4 - синаптическая щель, 5 - холинорецепторы на постсинап- тической мембране, 6 - складки на постсинаптической мембране
В ходе становления моторной иннервации существует этап, когда каждая миотрубка (мион) имеет множественную иннервацию. Однако позже множественная иннервация устраняется в процессе увеличивающейся двигательной активности. Особенностью ранних стадий развития мышечных волокон является разлитая чувствительность всей поверхности мембраны к ацетилхолину, присущая нервной стадии развития мышц. В процессе развития на каждом мионе сохраняется единственный синапс, формирование которого сопровождается появлением в постсинаптической мембране холинэстеразы. Увеличение ее концентрации приводит к повышению скорости гидролиза ацетилхолина. На постсинаптической мембране образуются складки, происходит концентрация холинорецепторов в синаптической зоне, в результате чего увеличивается амплитуда потенциала концевой пластинки (ПКП) и повышается надежность передачи возбуждения через синапс. Внесинаптическая поверхность мышечного волокна постепенно теряет чувствительность к ацетилхолину. После денервации она вновь распространяется на всю поверхность мембраны. Вследствие незрелости нервно-мышечного синапса у плода и новорожденного синаптическая передача возбуждения происходит относительно медленно: у взрослых длительность синаптической задержки около 0,5 мс, у новорожденных - около 4,5 мс. Соответственно низка и лабильность синапсов новорожденных. Без трансформации ритма через такой синапс передается не более 20 импульсов, а у взрослых – 100-150 импульсов в секунду. Таким образом, по мере созревания мионеврального синапса время перехода возбуждения с нерва на мышцу значительно (в среднем в 8 -10 раз) укорачивается и к 7 -8 годам жизни становится таким же, как у взрослых. Варианты организации тормозных процессов в ЦНС. Если рассмотреть «архитектуру» использования тормозных нейронов при организации нейронных сетей, цепей и рефлекторных дуг, то можно выделить ряд вариантов этой организации (это отражается в названии соответствующего вида торможения). 1. Реципрокное торможение (от лат. reciprocus - взаимный) было открыто английским физиологом Ч. Шеррингтоном и российским физиологом Н.Е. Введенским. Этот вид торможения основан на том, что одни и те же афферентные пути, через которые осуществляется возбуждение одной группы нервных клеток, обеспечивают через вставочные нейроны торможение других групп нейронов. Например, при возбуждении болевых рецепторов кожи конечности сигнал от ноцицепторов с участием афферентного нейрона поступает в спинной мозг, где переключается на альфа-мотонейрон мышц-сгибателей и одновременно на тормозной нейрон, который тормозит активность альфа-мотонейрона мышц-разгибателей. Реципрокное торможение характерно как для спинного мозга, так и для головного. Экспериментально доказано, что нормальное выполнение каждого естественного двигательного акта основано на использовании реципрокного торможения. 2. Возвратное, или антидромное, торможение наблюдается в отношении альфа-мотонейронов спинного мозга. При возбуждении альфа-мотонейрона нервный импульс направляется к мышечным волокнам, возбуждая их. Одновременно по коллатерали, идущей к тормозному нейрону (клетка Реншоу), импульс возбуждает эту тормозную клетку, которая в свою очередь вызывает торможение возбужденного ранее альфа-мотонейрона. Таким образом, альфа-мотонейрон, активируясь, через систему тормозного нейрона сам себя (возвратно, или антидромно) затормаживает. Чем выше активность альфа-мотонейрона, тем выраженнее тормозное влияние клеток Реншоу на этот нейрон (это проявляется в снижении частоты генерации потенциалов действия альфа-мотонейроном). 3. Латеральное торможение рассматривается как вариант возвратного торможения. Существует несколько видов использования в ЦНС возвратного торможения, в частности, латеральное торможение. Оно играет важную роль в организации обработки информации нейронами сетчатки. Суть его сводится к следующему. Под влиянием квантов света активируется фоторецептор. В свою очередь это вызывает изменение активности биполярной клетки, связанной с данным фоторецептором. Одновременно активируется рядом расположенный тормозной нейрон, который блокирует проведение возбуждения от соседнего фоторецептора к ганглиозной клетке. Тем самым происходит «вытормаживание» информации в соседних участках. Таким способом создаются условия для четкого видения предмета (две точки на сетчатке рассматриваются как две раздельные точки в том случае, если между ними есть невозбужденные участки). 4. Тоническое торможение отражает наличие постоянного тормозного влияния одной структуры на другую. Примером такого постоянного тормозного влияния являются тормозные нейроны коры больших полушарий, которые угнетают нейроны ретикулярной формации ствола мозга, нейроны таламуса и лимбической системы. 5. Общее центральное торможение открыто в 1937 И.С. Беритовым (Словарь физиологических терминов, 1987). Это нервный процесс, который развивается при любой рефлекторной деятельности. Он захватывает почти всю ЦНС, включая центры головного мозга. Такое торможение проявляется раньше, чем возникнет какая-либо двигательная реакция. Оказалось, что общее центральное торможение может проявляться при такой малой силе раздражителя, при которой двигательный эффект еще отсутствует. По мнению И.С. Беритова, такое торможение обеспечивает концентрацию возбуждения в определенных группах вставочных и двигательных нейронов, препятствуя возникновению других рефлекторных или поведенческих актов, которые могли бы возникнуть под влиянием раздражителей. Важную роль в формировании общего центрального торможения, вероятно, играет желатинозная субстанция спинного мозга. Не исключено, что общее центральное торможение играет важную роль в создании целостной поведенческой деятельности, а также в обеспечении избирательного возбуждения определенных рабочих органов. Пессимальное торможение как вариант вторичного торможения. Этот вид торможения развивается в возбуждающих синапсах в результате сильной и длительной деполяризации постсинаптической мембраны приходящими сюда высокочастотными импульсными потоками (подобно пессимуму Введенского в нервно-мышечном синапсе либо католической депрессии Вериго), что приводит к аккомодационным изменениям и снижению возбудимости (повышению порога возбуждения) в постсинаптической мембране, делающим ее неактивной. В частности, этот вид торможения лежит в основе пессимального торможения условнорефлекторной деятельности в коре больших полушарий. Торможение вслед за возбуждением как вариант вторичного торможения. Этот вид торможения развивается достаточно часто, так как возникает всякий раз на фоне следовой гиперполяризации мембраны нейрона после очередного его возбуждения. Для него характерен сравнительно кратковременный период существования, так как он определяется лабильностью нейрона, т.е. скоростью восстановления исходного уровня мембранного потенциала после генерации очередного потенциала действия. Свойства нервных центров Нервный центр - совокупность структур центральной нервной системы, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт. А. А. Ухтомский дал определение нервного центра как созвездие («констелляция») созвучно работающих нервных клеток. И сейчас это определение мало изменилось: принято говорить, что нервный центр - это совокупность нейронов, необходимых для осуществления определенного рефлекса или регуляции той или иной функции. О месте нахождения этих клеток судят по эффектам раздражения отдельных участков ЦНС, по нарушению функции после удаления тех или иных отделов мозга, в опытах с перерезками мозга на разных уровнях и др. Известно, что группы клеток, регулирующих одну и ту же функцию, могут располагаться в разных отделах ЦНС. При этом различно функциональное значение не только нейронов, лежащих в разных отделах ЦНС, но и нейронов одного и того же отдела. И. П. Павлов ввел понятие ядра и рассеянных элементов центра. При повреждении ядра центра происходит глубокое нарушение функции, а нарушение функции, возникающее при повреждении рассеянных элементов (нейронов расположенных к периферии от ядра), легко компенсируется. Расположение клеток одного и того же центра в разных отделах ЦНС ярко прослеживается на центре речи. Эту функцию обеспечивают нервные клетки, регулирующие двигательную активность речевых мышц языка и губ, мышц гортани, расположенные в продолговатом мозге. Высшая регуляция речевой функции осуществляется клетками, лежащими в височной, лобной и теменной областях коры больших полушарий. При повреждении теменной области человек теряет способность понимать речь, хотя и хорошо ее слышит. При повреждении лобных долей - нарушается двигательная речевая функция. Кроме того, одни и те же нейроны ЦНС могут участвовать в регуляции разных функций. Так, клетки центра глотания, расположенные в продолговатом мозге, обеспечивают последовательные сокращения мышц языка, неба, глотки, гортани и пищевода. Но поднятие мягкого неба происходит и при акте рвоты. Следовательно, нервные клетки, регулирующие движение мышц мягкого неба входят в состав и центра глотания и центра рвоты. Для нормального осуществления определенной функции необходима целостность всех отделов ЦНС, регулирующих эту функцию. В тех случаях, когда говорят о свойствах нервных центров, имеют в виду не такое широкое его определение, а лишь свойства нейронов, связанные с особенностями их строения и механизмом передачи возбуждения в синапсах. Нервные центры имеют ряд общих свойств, что во многом определяется структурой и функцией синаптических образований. 1. Одностороннее проведение возбуждения. В ЦНС - в ее нервных центрах, внутри рефлекторной дуги и нейронных цепей возбуждение, как правило, идет в одном направлении - от пресинаптической мембраны к постсинаптической, т.е. вдоль рефлекторной дуги от афферентного нейрона к эфферентному. Это связано со свойствами синапсов: для химических - с выработкой медиаторов в пресинаптической части синапса, диффузией их через синаптическую щель к хеморецепторам постсинаптической мембраны, а для большинства электрических синапсов - с полупроводниковыми свойствами их синаптических мембран. В целом это организует деятельность ЦНС и является одним из принципов координационной деятельности ЦНС. 2. Замедление проведения возбуждения в нервных центрах, или центральная задержка. Замедление проведения возбуждения по нервным центрам получило название центральной задержки. Она обусловлена медленным проведением нервных импульсов через синапсы, так как затрачивается время на следующие процессы: выделение медиатора из пресинаптических везикул, трансфузия его через синаптическую щель к постсинаптической мембране и генерация возбуждающего постсинаптического потенциала (ВПСП). При этом истинная синаптическая задержка (до начала генерации ВПСП) составляет 0,5 мс, а вместе со временем генерации ВПСП она достигает 1,5-2,5 мс. 3. Суммация возбуждения и суммация торможения. Принято выделять два вида суммации - временную и пространственную. Временная, или последовательная, суммация проявляется в том, что в области постсинаптической мембраны происходит суммация следов возбуждения во времени, т.е. на нейроне в области его аксонного холмика происходит интеграция событий, разыгрывающихся на отдельных участках мембраны нейрона на определенном отрезке времени. Например, если с определенным интервалом к нейрону 1 в точку А через возбуждающие синапсы приходят потенциалы действия (ПД) от возбуждающего нейрона 2, то на постсинаптической мембране нейрона 1 будут генерироваться ВПСП. Если эти ВПСП не достигают критического уровня деполяризации, то ПД на аксонном холмике не возникает. Если же частоту следования подпороговых импульсов от нейрона 1 увеличить, то на аксонном холмике может произойти суммация ВПСП, при которой суммарное изменение мембранного потенциала достигнет критического уровня деполяризации и нейрон 1 будет генерировать ПД, т.е. возбудится. Это явление носит название временной, или последовательной, суммации, так как в этом случае происходит суммация следов возбуждения во времени.
|
|||||||||
|
|
Последнее изменение этой страницы: 2019-11-02; просмотров: 235; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.200.143 (0.064 с.) |