Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Строение плазматической мембраны.
Согласно жидкостно-мозаичная модели, мембрана представлена бислоем фосфолипидных молекул. При этом гидрофобные концы молекул находятся внутри бислоя, а гидрофильные направлены в водную фазу, что способствует для образования раздела двух фаз: вне – и внутриклеточной. В фосфолипидном бислое интегрированы глобулярные белки, полярные участки которых образуют гидрофильную поверхность в водной фазе. Эти интегрированные белки выполняют различные функции: 1. Ионные каналы – транспортируют вещества внутрь клетки и обратно. 2. Выступают как переносчики определенных молекул через мембрану. 3. Являются ферментами и катализируют ассоциированные с мембраной реакции. 4. Выполняют адгезивную роль, связывая цитоскелет с внеклеточным матриксом. 5. Служат в качестве мембранных рецепторов для получения и преобразования химических сигналов из внешней среды. 6. Идентифицируют вещества, вступающие в контакт с мембраной клетки. Общее представление о структуре и функциях ионных каналов. Ионные каналы – особые образования в мембране клетки, представляющие собой олигомерные (состоящие из нескольких субъединиц) белки. Центральным образованием канала является молекула белка, которая пронизывает мембрану таким образом, что в ее гидрофильном центре формируется канал-пора, через которую в клетку способны проникать соединения, диаметр которых не превышает диаметра поры (обычно- это ионы). Вокруг главной субъединицы канала располагается система из нескольких субъединиц, которые формируют участки для взаимодействия с мембранными регуляторными белками, различными медиаторами, а также фармакологически активными веществами. Классификация ионных каналов по их функциям: 1) по количеству ионов, для которых канал проницаем, каналы делят на селективные (проницаемы только для одного вида ионов) и неселективные (проницаемы для нескольких видов ионов). Селективные по характеру ионов, которые они пропускают на Na+, Ca++, Cl-, K+-каналы; 2) по способу регуляции делятся на потенциалзависимые (электровозбудимые, потенциалуправляемые), потенциалнезависимые (хемовозбудимые, лиганд-рецептор -зависимые, хемоуправляемые) и механовозбудимые (механоуправляемые). Потенциалзависимые каналы реагируют на изменение потенциала мембраны клетки, и при достижении потенциалом определенной величины, канал переходит в активное состояние, начиная пропускать ионы по их градиенту концентрации.
Потенциалнезависимые каналы реагируют не на изменение мембранного потенциала, а на взаимодействие рецепторов, с которыми они взаимосвязаны, и их лигандов. Так, Cl--каналы связаны с рецепторами g-аминомасляной кислоты и при взаимодействии этих рецепторов с ней они активируются и обеспечивают ток ионов хлора в клетку, вызывая ее гиперполяризацию и снижение возбудимости. Состояние проницаемости механовозбудимыех каналов изменяется при механических воздействиях на мембрану, вызывающих нарушение структурной упаковки молекул в мембране и ее растяжение. Эти каналы широко представлены в цитоплазматической мембране механорецепторов сосудов, внутренних органов, кожи, ППМ и ГМ.
Строение ионного канала.
Различают следующие основные виды электрических ответов возбудимых клеток: 1. Локальные потенциалы (рецепторный, потенциал, локальный ответ, ВПСП (возбуждающий постсинаптический птенциал), ТПСП (тормозной постсинаптический потенциал) Мембранный потенциал покоя (МПП) Потенциал действия (ПД). Мембранный потенциал покоя и его происхождение. Термином «мембранный потенциал покоя » принято называть трансмембранную разность потенциалов, существующую между цитоплазмой и окружающим клетку наружным раствором. Когда клетка (волокно) находится в состоянии физиологического покоя, ее внутренний заряд отрицателен по отношению к наружному, условно принимаемому за нуль. У разных тканей мембранный потенциал характеризуется разной величиной: самый большой у мышечной ткани -80 -90 мВ, у нервной -70 мВ, у соединительной -35 -40 мВ, у эпителиальной -20мВ.
Образование МПП зависит от концентрации ионов К+, Nа+, Са2+, Сl-, и от особенностей строение мембраны клетки. В частности важным в образовании МПП является то, что мембранные ионные каналы, обладают свойствами: 1. Селективностью (избирательной проницаемостью) 2. Электровозбудимостью.
В состоянии покоя натриевые каналы все закрыты, а большинство калиевых – открыты. Каналы могут открываться и закрываться. В мембране существуют каналы утечки (неспецифические), которые проницаемы для всех элементов, но более проницаемы для калия. Ионы движутся через эти каналы по концентрационному и электрохимическому градиенту. Согласно мембранно-ионной теории наличие МПП обусловлено: Ø непрерывным движением ионов по ионным каналам мембраны, Ø постоянно существующей разностью концентраций катионов по обе стороны мембраны, Ø непрерывной работой натрий-калиевого насоса. Ø различной проницаемостью каналов для этих ионов. В состоянии покоя мембрана клетки проницаема только для ионов К+. Ионы калия в состоянии покоя постоянно выходят в окружающую среду, где высокая концентрация Nа+. Поэтому, в состоянии покоя, наружная поверхность мембраны заряжена положительно. Высокомолекулярные органические анионы (белки) концентрируются у внутренней поверхности мембраны и определяют ее отрицательный заряд. Они же электростатически удерживают ионы К+ с другой стороны мембраны. Основную роль в образовании МПП принадлежит ионам К+. Блокируют проницаемость К+ тетраэтиламмония, аминопиридины. Несмотря на потоки ионов через каналы утечки разность концентрации ионов не выравнивается, т.е. сохраняется всегда постоянной. Этого не происходит потому, что в мембранах существуют Nа+ - К+- насосы. Они непрерывно откачивают Nа+ из клетки и против градиента концентрации вводят в цитоплазму К+. На 3 иона Nа+, которые выводятся из клетки, внутрь вводится 2 иона К+. Перенос ионов против градиента концентрации осуществляется активным транспортом (с затратой энергии). В случае отсутствия энергии АТФ клетка погибает. Наличие потенциала покоя позволяет клетке практически мгновенно после действия раздражителя перейти из состояния функционального покоя в состояние возбуждения. При возбуждении происходит снижение величины исходного потенциала покоя с перезарядкой мембраны. Когда внутренний заряд мембраны становится менее отрицательным наступает деполяризация мембраны и начинает развиваться потенциал действия, увеличение отрицательного – гиперполяризация (возбудимость снижается). Потенциал действия и механизм его происхождения. Соотношение фаз возбудимости с фазами потенциала действия Потенциалом действия называют быстрое колебание мембранного потенциала, возникающее при действии раздражителя пороговой силы в нервных и мышечных клетках. В его основе лежат изменения ионной проницаемости мембраны. Амплитуда и характер изменений потенциала действия мало зависят от силы вызывающего его раздражителя, важно лишь, чтобы эта сила была не меньше пороговой. Возникнув, ПД распространяется вдоль мембраны, не изменяя своей амплитуды. В нем различают фазы: 1) Деполяризации: а) медленная деполяризация; б) быстрая деполяризация. 2) Реполяризация: а) быстрая реполяризация; б) медленная реполяризация (отрицательный следовой потенциал) 3) Гиперполяризация (положительный следовой потенциал)
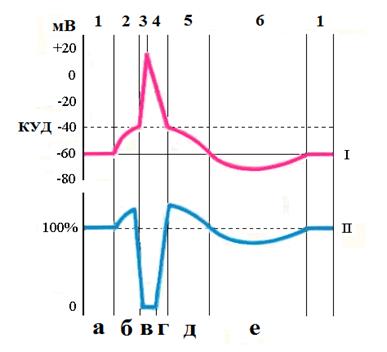
Возникновение потенциала действия (деполяризация)
I – мембранный потенциал 1 - потенциал покоя, 2-- медленная деполяризация, 3 - быстрая деполяризация, 4 - быстрая реполяризация, 5 – медленная реполяризация, 6- гиперполяризация
II - возбудимость а – нормальная, б – повышенная, в –абсолютная рефрактерность, г – относительная рефрактерность, д – супернормальность, е -субнормальность Механизм возникновения ПД. Деполяризация После действия раздражителя пороговой силы потенциал покоя уменьшается, повышает проницаемость каналов для ионов натрия, который входит в клетку. Поток ионов натрия в клетку выше, чем поток ионов К+ из клетки. Поступление в клетку иона натрия уменьшает электроотрицательность внутренней поверхности мембраны, что способствует активации новых ионных натриевых каналов и дальнейшему поступлению в клетку иона натрия. Внутренняя сторона мембраны становится положительной, а на поверхности ионы Cl- образуют отрицательный заряд. Перезарядка мембраны, или овершут, характерен для большинства возбудимых клеток. Амплитуда овершута характеризует состояние мембраны и зависит от состава вне- и внутриклеточной среды. Смена зарядов на внутренней и наружной поверхности мембраны соответствует фазе деполяризации. Реполяризация После смены зарядов на наружней и внутренней поверхности клеточной мембраны происходит инактивации (закрытия) натриевых каналов. Поступление в клетку ионов натрия становится минимальным. Выход из клетки ионов калия восстанавливает электроотрицательность внутренней поверхности мембраны, что соответствует фазе реполяризации. В последующем натрий/калиевый насос мембраны удаляет из клетки поступивший при деполяризации натрий и восстанавливает исходную концентрацию калия, который вышел из клетки при реполяризации. Основная роль в образовании ПД принадлежит ионам Na+. Блокирует транспорт нартия тетродотоксин.
|
|||||||
|
|
Последнее изменение этой страницы: 2019-08-19; просмотров: 242; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.217.6.114 (0.015 с.) |