Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Структурно-функциональные особенности синтетического аппарата клетки
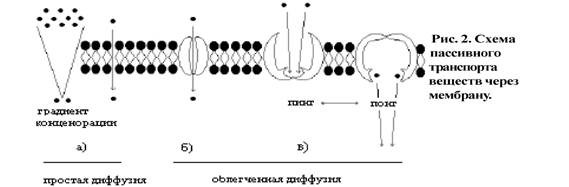
Клетка - целостная элементарная система, способная к самовоспроизведению и саморегуляции метаболических процессов. Эукариотическая клетка состоит из 3-х частей: поверхностного аппарата, цитоплазмы и ядра. II. Поверхностный аппарат клетки (рис. 1) состоит из плазмалеммы (1), надмембраннного (2) и субмембранного комплексов (3).
Плазмалемма (плазматическая мембрана) образована белками (-60%) и липидами (-40%). В состав липидов входят гликолипиды, стеролы, но наиболее распространены фосфолипиды. Фосфолипиды состоят из гидрофильной головки (глицерин) и гидрофобного хвоста (жирные кислоты); данная полярность липидов и позволяет образовывать в водной среде билипидный слой. По расположению в плазмалемме выделяют три вида белков: интегрГальные (а), полуинтегральные (б), периферические (в). "Белки как айсберги плавают в липидном море" поодиночке, но чаще сцепленными группами. Надмембранный комплекс (2), выполнен гликолипидами и гликопептидами, в животной клетке представлен гликокаликсом, в растительной - клеточной стенкой. Субмембранный слой (3) образуется путем скопления микротрубочек и микрофиламентов цитоскелета под плазмалеммой. Функции поверхностного аппарата клетки: барьерная (защитная, пограничная), обменно-транспортная, рецепторная, метаболическая, контактная (в многоклеточном организме).
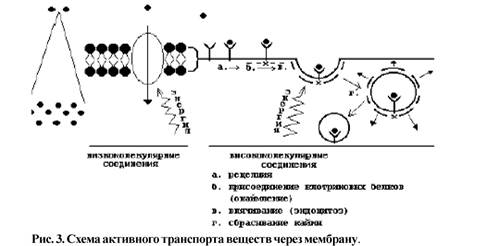
Активный транспорт (рис.З) идет против градиента концентрации веществ, с затратой энергии. Активный транспорт можно разделить на 2 вида: активный транспорт низкомолекулярных соединений (рис.З-1) /Пример: (Na++K+)-Hacoc/ и активный транспорт высокомолекулярных соединений (транспорт веществ в клетку
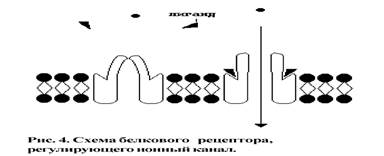
Рецепторная функция поверхностного аппарата связана с идентификацией информационного вещества - лиганда (как правило, это гормон) при помощи рецептора и адекватного ответа на данный стимул (это может быть запуск химического процесса в клетке или открытие транспортного канала) (рис.4).
Существуют и другие органоиды, имеющие свое специфическое строение и функции.
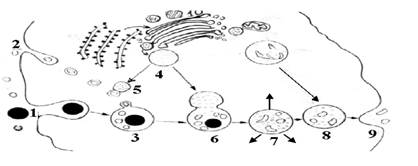
Рис. 7. Схематическое изображение литической системы клетки (пример мембранного конвейера). Литический цикл можно разделить на три основные этапа: 1. Образование эндосомы (в результате эндоцитоза) или аутосомы (в результате
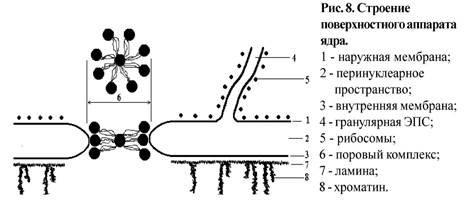
Кариоплазма (ядерный сок) - внутренняя бесструктурная среда ядра, по своим физико-химическим свойствам это белковый коллоид, в котором располагаются все структуры ядра. Основу ядерного матрикса составляют фибриллярные белки, создающие "скелет" ядра или участвующие в репликации и транскрипции.
Структурная основа мембран Рассматривая с помощью электронного микроскопа ультратонкий срез клетки, можно увидеть огромное количество тонких двойных линий толщиной от 7 до 10 нм, которые представляют собой срез через биологические мембраны. Изучение структуры и функционирования биологических мембран – одна из наиболее быстро развивающихся областей биофизики. Эта область имеет важное значение и для теоретической, и для клинической медицины. Многие патологические процессы в клетке, включая такие, как раковое перерождение, нарушение функционирования при атеросклерозе, нарушения диеты или тканевая гипоксия, повреждение при вирусных и бактериальных инфекциях, а также при интоксикациях и действии УФ и ионизирующей радиации, — прямо или косвенно связаны с нарушением барьерной и структурной функции мембран. Действие многих лекарств направлено на изменение функции мембран, а эффективность лекарственных соединений зависит от их способности проникать через мембраны или связываться с ними.
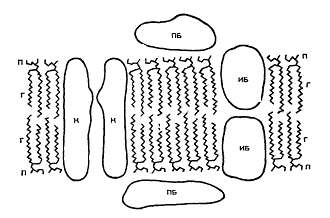
Рис. 6. Общая схема строения биологических мембран. Мембраны состоят из двойного слоя фосфолипидных молекул, каждая из которых имеет полярную головку (П) и неполярный, гидрофобный «хвост» (Г). ПВ — периферические белки, ИВ — интегральные белки. Пронизывая бислой, ИБ могут образовывать канал (пору) для ионов (К). Структурную основу любой биологической мембраны (рис. 6.) как целого составляет фосфолипидный бимолекулярный слой; именно он выполняет в мембране функцию барьера для ионов и водорастворимых молекул и функцию основы, матрицы для мембранных ферментов, рецепторов и других встроенных в мембраны белков, гликолипидов и гликопротеидов. Белки мембран могут либо находиться на поверхности липидного слоя, удерживаемые преимущественно электростатическими силами (периферические белки), либо проникать глубоко в липидный бислой или даже пронизывать его насквозь; в этом случае они связаны с липидами прочными гидрофобными взаимодействиями (собственные или интегральные белки). В плазматических мембранах большинства клеток к липидному бислою примыкает гликока–ликс, состоящий из гликолипидов и гликопротеидов; в составе липидов плазматических мембран много холестерина.
Молекулы фосфолипидов, входящие в состав биологических мембран, обладают свойствами амфифильности, которое заключается в том, что часть молекулы полярна (а потому гидрофильна), а другая — неполярна (и гидрофобна). Это обусловлено особенностями химической структуры фосфолипидов, молекулы которых строятся из глицерина, жирных кислот, фосфорной кислоты и полярного соединения, характерного для каждого класса фосфолипидов (холин, этаноламин, серин и др.). По своей форме молекулы фосфолипидов напоминают сплющенный цилиндр, около 1/4 которого (по длине) гидрофильна, а 3/4 — гидрофобна (рис. 7.).
Рис. 7. Структура молекулы фосфатидилхолина (лецитина) Мембранные белки К мембранным белкам относятся белки, которые встроены в клеточную мембрану или мембрану клеточной органеллы или ассоциированы с таковой. Около 25 % всех белков являются мембранными Мембранные белки могут быть классифицированы по топологическому или биохимическому принципу. Топологическая классификация основана на том, сколько раз белок пересекает липидный бислой. В соответствии с этим критерием белки подразделяются на монотопические, битопические и политопические: монотопические белки взаимодействуют с одной поверхностью мембраны и не пересекают её; битопические пронизывают мембрану насквозь и взаимодействуют с обеими её поверхностями; политопические пронизывают мембрану несколько раз (многократное взаимодействие с липидами). Понятно, что первые относятся к периферическим белкам, а вторые и третьи к интегральным. Топологическая классификация[править | править вики-текст] По отношению к мембране мембранные белки делятся на поли- и монотопические. Политопические, или трансмембранные, белки полностью пронизывают мембрану и, таким образом, взаимодействуют с обеими сторонами липидного бислоя. Как правило, трансмембранный фрагмент белка является альфа-спиралью, состоящей из гидрофобных аминокислот (возможно от 1 до 20 таких фрагментов). Только у бактерий, а также в митохондриях и хлоропластах трансмембранные фрагменты могут быть организованы как бета-складчатая структура (от 8 до 22 поворотов полипептидной цепи). Интегральные монотопические белки постоянно встроены в липидный бислой, но соединены с мембраной только на одной стороне, не проникая на противоположную сторону. Биохимическая классификация[править | править вики-текст] По биохимической классификации мембранные белки делятся на интегральные и периферические. Интегральные мембранные белки прочно встроены в мембрану и могут быть извлечены из липидного окружения только с помощью детергентов или неполярных растворителей. По отношению к липидному бислою интегральные белки могут быть трансмембранными политопическими или интегральными монотопическими. Периферические мембранные белки являются монотопическими белками. Они либо связаны слабыми связями с липидной мембраной, либо ассоциируют с интегральными белками за счёт гидрофобных, электростатических или других нековалентных сил. Таким образом, в отличие от интегральных белков они диссоциируют от мембраны при обработке соответствующим водным раствором (например, с низким или высоким pH, с высокой концентрацией соли или под действием хаотропного агента). Эта диссоциация не требует разрушения мембраны. Мембранные белки могут быть встроены в мембрану за счёт жирнокислотных или пренильных остатков либо гликозилфосфатидилинозитола, присоединённых к белку в процессе их посттрансляционной модификации. Гликокаликс Гликокаликс - это обогащенная углеводами периферическая зона на поверхности большинства эукариотических клеток (клеточная оболочка). При использовании различных красителей, например рутениевого красного, эту зону можно отчетливо видеть в электронном микроскопе. Гликокаликс — «заякоренные» в плазмалемме молекулы олигосахаридов, полисахаридов, гликопротеинов и гликолипидов. Гликокаликс выполняет рецепторную и маркерную функции, а также участвует в обеспечении избирательности транспорта веществ и пристеночном (примембранном) пищеварении Функции плазмалеммы Плазмолемма отмечается определенной асимметричностью. Все вуглеводмистки части гликопротеидных и гликолипидного молекул мембраны выступают на ее внешнюю поверхность и участвуют в формировании надмембранный слоя. Внешняя поверхность содержит различные молекулы, которые называют рецепторами. Холестерин находится более во внутреннем слое мембраны. Выявлено также, что белки, содержащиеся на внешний и внутренней поверхности мембраны, идентичны. Сложное строение плазмолеммы обеспечивает выполнение ею важных функций. Мембранные рецепторы взаимодействуют с молекулами клеточного окружения, влияющего на активность клетки, ее стимулирования или подавления. а) Барьерная функция - разграничение содержимого клетки от ее окружения. Благодаря этой функции обеспечивается характерная структура клеточной поверхности и защита от случайного проникновения веществ в клетку. б) Транспортная функция Мембранный транспорт веществ может осуществляться как однонаправленное перенос молекул определенного вещества или общий транспорт двух различных молекул в одном или противоположном направлениях. Мембранный транспорт бывает трех видов: пассивный, активный и транспорт в мембранной упаковке. Пассивный транспорт - это простая и облегченная диффузия (от лат. Diffusio - распространение, распространение, растекание). Простая (нейтральная) диффузия обеспечивает пропускание мелких молекул (О2, Н2О, СО2) со скоростью, пропорциональной градиенту концентрации с обеих сторон мембраны. Это переход ионов или молекул, вызванный их бровнивським движением, через мембраны из зоны, где эти вещества находятся в большей концентрации, в зону с более низкой концентрацией до тех пор, пока концентрации по обе стороны мембраны выровняются. При простой диффузии незаряженные вещества проходят между липидными молекулами мембраны или через каналы, сформированные белками. Облегченная диффузия осуществляется через ионные каналы или белки - переносчики, которые обладают специфичностью по отношению к транспортируемых молекул. Белки - переносчики - это специфические белки, которые связывают вещество и переносят ее через мембрану. Роль ионных каналов выполняют трансмембранные белки, которые образуют мелкие водные поры. Через них по электрохимическому градиенту транспортируются мелкие водорастворимые молекулы и ионы. Эти каналы состоят из транспортной системы и воротной механизма, который открывает канал на некоторое время на: (1) изменения мембранного потенциала, (2) механические воздействия (например, в волосковых клетках внутреннего уха), (3) на связывание лиганда (сигнальной молекулы или иона). Белки - переносчики также трансмембранными белками, обеспечивающих транспорт специфических белков через плазмолемму. Они участвуют в механизмах как пассивного, так и активного транспорта. Благодаря наличию в плазмолемме мембранных транспортных белков, специфичных для каждого соединения, большие незаряженные полярные молекулы диффундируют легко. Эти белки могут функционировать по принципу унипорта (перенос одного вещества через мембрану) или котранспорта (перенос двух веществ). Последний может быть в виде симпорта (перенос двух веществ в одном направлении) или антипорта (перенос двух веществ в противоположных направлениях). Активный транспорт. Как уже отмечалось, диффузия происходит только по градиенту концентрации, облегченная диффузия - только с участием ионных каналов, тогда как активный транспорт осуществляется против градиента и требует участия ионов металлов. Перенос глюкозы, аминокислот и жирных кислот, а также некоторых ионов из тканевой жидкости в цитоплазму осуществляется с помощью специальных ферментов с использованием энергии. Такие системы, которые используют энергию и действуют с помощью ферментов, иногда называются системами активного переноса. Натриево - калиевый канал и натриево - калиевая помпа - это система, которая обеспечивает перемещение ионов натрия и калия через плазмолемму. Перенос ионов осуществляет специальный фермент - переносчик Na + - K + - АТФ -аза, который использует энергию клетки и откачивают ионы натрия через плазмолемму наружу, одновременно он захватывает ионы калия извне и освобождает их внутри клетки. Таким образом удерживается разница концентрации натрия и калия: концентрация натрия выше в тканевой жидкости (снаружи плазмолеммы), а калия - в цитоплазме. При гидролизе одной молекулы АТФ из клетки випомповуються 3 ионы Na + i 2 ioны K + вводятся в нее. В то же время белок - переносчик иона Na + транспортирует глюкозу в клетку. Натриево - калиевая помпа обеспечивает поддержание постоянного объема клетки (путем регулирования осмотического давления), а также мембранного потенциала. Транспорт в мембранной упаковке характерен тем, что вещества, которые переносятся через плазмолемму, обычно окружены мембраной (рис. 2.17). Так, транспорт веществ в клетку называют эндоцитоза, а из клетки - экзоцитоза. Капля выделяемой может быть окружена мембраной (например, липосома), или нет. При эндоцитозе (от греч. Еndo - внутрь и cytos - клетка) материал, который находится во внеклеточном пространстве, будучи охваченным складками плазмолеммы, попадает в клетку в виде эндоцитозных пузырька, или эндосомы (рис. 2.17). Далее содержимое эндосомы подвергается процессингу (внутриклеточной обработке). Разновидностями эндоцитоза является фагоцитоз (от греч. Рhagein - поедать и cytos - клетка) - захвата и поглощения крупных (до 1 мкм) и плотных частиц, и пиноцитоза (от греч. Pinein - пить и cytos - клетка) - втягивание окруженных плазмолеммой жидкостей и растворимых веществ. Рецепторно -опосредованный эндоцитоз значительно эффективнее, так как он опосредован рецепторами, которые связываются с молекулами фагоцитированные объекта - лигандами (от лат. Ligare - связывать). После поглощения вещества комплекс рецептор - лиганд расщепляется, и рецепторы могут вновь возвращаться в плазмолемму. Примером такого рецепторно опосредованного эндоцитоза может служить фагоцитоз лейкоцитов микроба. На плазмолемме лейкоцита есть рецепторы к иммуноглобулинам (антитела), и если поверхность микробов покрыта антителами - опсонинами (от греч. Оpson - приправа), то скорость фагоцитоза резко возрастает. Особое усиление приобретает эндоцитоз в окаймленных пузырьков, где в местах их формирования накапливаются рецепторы и особые белки - клатрины, связывающие лиганды. Когда пузырек втягивается в цитоплазму, его содержание очень быстро подлежит процессинга - протеолиза. Если клатринова оболочка не теряется, то его содержание остается неизменным. Экзоцитоз (от греч. Еxo - наружу и cytos - клетка) - процесс, при котором мембранные екзоцитозни пузырьки, сформировавшиеся в цитоплазме, приближаются к плазмолеммы и сливаются с ней своей мембраной, которая встраивается в плазмолемму. При этом содержание пузырька выделяется во внеклеточное пространство. Трансцитоза (от лат. Trans - через и cytos - клетка) - процесс, при котором на одной поверхности клетки формируется эндоцитозных пузырек, который переносится на противоположную поверхность клетки как екзоцитозний и выделяет свое содержимое во внеклеточное пространство. Трансцитоза характерен для клеток с периферической тонкой плазмолеммой (эндотелиоциты), которые выстилают мелкие кровеносные сосуды. В этих клетках пузырьки, сливаясь, могут образовывать временные трансцелюлярни каналы, через которые транспортируются водорастворимые вещества. В процессах как эндоцитоза, так и экзоцитоза, важную роль имеют особые фузогенни (от лат. Fusio - слияние) мембранные белки, которые концентрируются в участках инвагинации (впячивания) плазмолеммы. При эндоцитозе они способствуют смыканию складок плазмолеммы, а во время экзоцитоза обеспечивают встраивания мембран пузырька в плазмолемму (рис. 2.17). Сбалансированность процессов эндоцитоза и экзоцитоза достигается таким образом, что при эндоцитозе пузырьки формируются из плазмолеммы и ее поверхность должна уменьшаться, тогда как при экзоцитоза мембраны пузырьков встраиваются в плазмолемму и увеличивают ее площадь. Однако активные процессы экзо - и эндоцитоза не влияют существенно на площадь поверхности плазмолеммы в связи с уравновешиванием этих процессов. Постоянный круговорот мембран в клетке получил название " мембранного конвейера ". Выведение веществ из клетки может осуществляться в виде: (1) секреции (от лат. Sekretio - выделение) - выделение нужных организму веществ, например, слюнные железы вырабатывают слюну, молочные - молоко; (2) экскреции - удаление ненужных или вредных веществ, например, мочевина, мочевая кислота удаляются почками; (3) рекреции (цитопемпсис) - транспорта, при котором клетка поглощает вещества одной поверхностью, а выводит их без изменений противоположной (например, минеральные соли, некоторые лекарства). в) Рецепторная функция плазмолеммы - это восприятие клеткой химических сигналов с ее микроокружения. Осуществляется она в основном с участием специальных сложных рецепторных белков плазмолеммы, содержащие углеводный компонент. Дальнейшая передача сигналов внутрь клетки в значительной степени осуществляется с помощью аденилатциклазной системы. Рецепторная функция плазмолеммы определяет взаимоотношения клеток с окружающей средой и с соседними клетками. Она осуществляется путем распознавания данной клеткой других клеток и прикрепления к ним; распознавания межклеточного вещества и прикрепления к ее элементам (базальной мембраны, волокон соединительной ткани); взаимодействия с сигнальными молекулами (гормонами, медиаторами, цитокинами и т.п.). Рецепторы обеспечивают различные таксисы (реотаксис, хемотаксис). Рецепторная функция лежит в основе таких явлений, как оплодотворение, перемещения клеток, ответ на гормональные влияния. Считают, что каждая клетка имеет рецепторы к инсулину, существуют рецепторы к лецитина, дофамина и др.. Благодаря разграничительной функции клетка может сохранять свою индивидуальность, тогда как транспортная функция чрезвычайно важна для его жизни и деятельности. Сочетание этих функций обеспечивает гомеостаз клетки - поддержание постоянного состава внутренней среды клетки.
|
|||||||||
|
|
Последнее изменение этой страницы: 2019-05-20; просмотров: 498; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.218.168.16 (0.05 с.) |