Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Морфофункциональная характеристика поперечнополосатых мышечных тканей сердечного типа
Сердечная мышечная ткань представлена специфической формой поперечнополосатых мышц. По сравнению со скелетными мышцами, она имеет ряд особенностей. В отличие от краевого расположения ядер в клетке скелетных мышц, ядра в клетке мышечной ткани сердца располагаются в центре клетки. Сами клетки по диаметру меньше мышечных волокон скелетных мышц. В противоположность мышечным волокнам скелетных мышц, которые снаружи не имеют фибриллярных структур, необходимых для связывания между собой, клетки мышечной ткани сердца связаны друг с другом особыми вставочными дисками. Такая организация мышечных клеток сердца дает возможность электрическому импульсу веерообразно распространяться по стенкам обоих предсердий и внутренней поверхности желудочков. Еще одна особенность сердечной мышцы заключается в способности некоторых ее клеток генерировать импульсы не только в ответ на внешние раздражители, но и спонтанно. Активность клеток мышцы сердца находится под контролем автономной нервной системы. Сердечная мышечная ткань Гистогенез и виды клеток. Источники развития сердечной поперечнополосатой мышечной ткани — симметричные участки висцерального листка спланхнотома в шейной части зародыша — так называемые миоэпикардиалъные пластинки. Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза возникает 3 вида кардиомиоцитов: 1. рабочие, или типичные, или же сократительные, кардиомиоциты, 2. атипичные кардиомиоциты (сюда входят пейсмекерные, проводящие и переходные кардиомиоциты, а также 3. секреторные кардиомиоциты. Рабочие (сократительные) кардиомиоциты образуют свои цепочки. Укорачиваясь, они обеспечивают силу сокращения всей сердечной мышцы. Рабочие кардиомиоциты способны передавать управляющие сигналы друг другу. Синусные (пейсмекерные) кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления. Они воспринимают управляющие сигналы от нервных волокон, в ответ на что изменяют ритм сократительной деятельности. Синусные (пейсмекерные) кардиомиоциты передают управляющие сигналы переходным кардиомиоцитам, а последние — проводящим. Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее — другим проводящим кардиомиоцитам. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим.
Секреторные кардиомиоциты выполняют особую функцию. Они вырабатывают гормон - натрийуретический фактор, участвующий в процессах регуляции мочеобразования и в некоторых других процессах. Сократительные кардиомиоциты имеют удлиненную (100—150 мкм) форму, близкую к цилиндрической. Их концы соединяются друг с другом, так что цепочки клеток составляют так называемые функциональные волокна (толщиной до 20 мкм). В области контактов клеток образуются так называемые вставочные диски. Кардиомиоциты могут ветвиться и образуют трехмерную сеть. Их поверхности покрыты базальной мембраной, в которую снаружи вплетаются ретикулярные и коллагеновые волокна. Ядро кардиомиоцита (иногда их два) овальное и лежит в центральной части клетки. У полюсов ядра сосредоточены немногочисленные органеллы общего значения. Миофибриллы слабо обособлены друг от друга, могут расщепляться. Их строение аналогично строению миофибрилл миосимпласта скелетного мышечного волокна. От поверхности плазмолеммы в глубь кардиомиоцита направлены Т-трубочки, находящиеся на уровне Z-линии. Их мембраны сближены, контактируют с мембранами гладкой эндоплазматической (т.е. саркоплазматической) сети. Петли последней вытянуты вдоль поверхности миофибрилл и имеют латеральные утолщения (L-системы), формирующие вместе с Т-трубочками триады или диады. В цитоплазме имеются включения гликогена и липидов, особенно много включений миоглобина. Механизм сокращения кардиомиоцитов такой же, как у миосимпласта. Кардиомиоциты соединяются друг с другом своими торцевыми концами. Здесь образуются так называемые вставочные диски: эти участки выглядят как тонкие пластинки при увеличении светового микроскопа. Фактически же концы кардиомиоцитов имеют неровную поверхность, поэтому выступы одной клетки входят во впадины другой. Поперечные участки выступов соседних клеток соединены друг с другом интердигитациями и десмосомами. К каждой десмосоме со стороны цитоплазмы подходит миофибрилла, закрепляющаяся концом в десмоплакиновом комплексе. Таким образом, при сокращении тяга одного кардиомиоцита передается другому. Боковые поверхности выступов кардиомиоцитов объединяются нексусами (или щелевыми соединениями). Это создает между ними метаболические связи и обеспечивает синхронность сокращений.
Возможности регенерации сердечной мышечной ткани. При длительной усиленной работе (например, в условиях постоянно повышенного артериального давления крови) происходит рабочая гипертрофия кардиомиоцитов. Стволовых клеток или клеток-предшественников в сердечной мышечной ткани не обнаружено, поэтому погибающие кардиомиоциты (в частности, при инфаркте миокарда) не восстанавливаются, а замещаются элементами соединительной ткани.
Строение и функции ядра Клеточное ядро (см. рис. 1 и 2) имеет важнейшее значение в жизнедеятельности клетки, поскольку служит хранилищем наследственной информации, содержащейся в хромосомах (см. ниже). Ядро есть в любой эукариотической клетке.
Рис. 1. Строение клетки эукариот. Обобщенная схема
Рис. 2. Строение клетки по данным электронной микроскопии Ядро ограничено ядерной оболочкой, отделяющей его содержимое (кариоплазму) от цитоплазмы. Оболочка состоит из двух мембран, разделенных промежутком. Обе они пронизаны многочисленными порами, благодаря которым возможен обмен веществами между ядром и цитоплазмой. В ядре клетки у большинства эукариот находится от 1 до 7 ядрышек. С ними связаны процессы синтеза РНК и тРНК. Основные компоненты ядра - хромосомы, образованные из молекулы ДНК и различных белков. В световом микроскопе они хорошо различимы лишь в период клеточного деления (митоза, мейоза). В неделящейся клетке хромосомы имеют вид длинных тонких нитей, распределенных по всему объему ядра. Во время деления клеток хромосомные нити образуют плотные спирали, вследствие чего становятся видимыми (с помощью обычного микроскопа) в форме палочек, «шпилек». Весь объем генетической информации распределен между хромосомами ядра. В процессе их изучения были выявлены следующие закономерности: в ядрах соматических клеток (т. е. клеток тела, неполовых) у всех особей одного вида содержится одинаковое количество хромосом, составляющих набор хромосом (рис. 3); для каждого вида характерен свой хромосомный набор по их количеству (например, у человека 46 хромосом, у мушки дрозофилы — 8, у аскариды — 4, у речного рака — 196, у лошади — 66, у кукурузы — 104); хромосомы в ядрах соматических клеток могут быть сгруппированы парами, получившими название гомологичных хромосом на основании их сходства (по строению и функциям); в ядрах половых клеток (гамет) из каждой пары гомологичных хромосом содержится только одна, т. е. общий набор хромосом вдвое меньше, чем в соматических клетках; одинарный набор хромосом в половых клетках называется гаплоидным и обозначается буквой n, а в соматических - диплоидным (2n).
Рис. 3. Хромосомы разных видов растений и животных, изображенные в одном масштабе: 1,2 — амеба; 3, 4 — диатомовые водоросли; 5-8, 18, 19 — зеленые водоросли; 9 — мухомор; 10 — липа; 11-12 — дрозофила; 13 — семга; 14 — скерда (семейство сложноцветных); 15 — растение из семейства ароидных; 16 — бабочка-хохлатка; 17 — насекомое из семейства саранчовых; 20 — клоп-водомерка; 21 — цветочный клоп; 22 — земноводное амбистома; 23 — алоэ (семейство лилейных)
Из изложенного ясно, что каждая пара гомологичных хромосом образована объединением отцовских и материнских хромосом при оплодотворении, т. е. слиянии половых клеток (гамет). И наоборот, при образовании половых клеток из каждой пары гомологичных хромосом в гамету попадает только одна. Хромосомы разных гомологичных пар отличаются по размерам и форме (рис. 4 и 5).
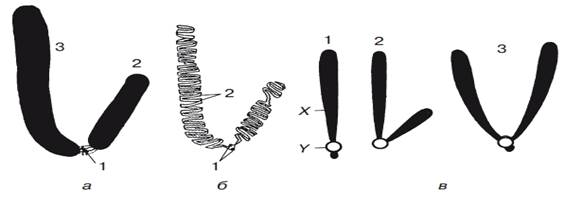
Рис. 4. Строение и типы хромосом:? — внешний вид (1 — цетромера; 2 — короткое плечо; 3 — длинное плечо); ? — внутренняя структура той же хромосомы (1 — центромера; 2 — молекулы ДНК);? — типы хромосом (1 — одноплечая; 2 — разноплечая; 3 — равноплечая: X — плечо, Y — центромера)
Рис. 5. Хромосома состоит из ДНК и белков. Молекула ДНК реплицируется. Две идентичные двойные спирали ДНК остаются соединенными в области центромеры. Эти копии превращаются в отдельные хромосомы позднее, во время деления клетки В теле хромосом выделяют первичную перетяжку (называемую центромерой), к которой прикрепляются нити веретена деления. Она делит хромосому на два плеча. Хромосомы могут быть равноплечими, разноплечими и одноплечими.
|
|||||||
|
|
Последнее изменение этой страницы: 2019-05-20; просмотров: 399; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.142.171.180 (0.009 с.) |