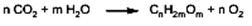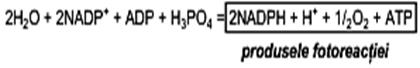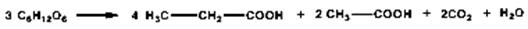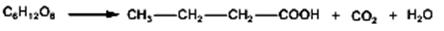Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
III.1.1. Biosinteza (anabolismul) glucidelor
Biosinteza monoglucidelor Biosinteza monoglucidelor poate fi exprimată printr-o reacţie globală, pornind de la CO2 şi H2O, substanţele precursoare, practic inepuizabile în natură:
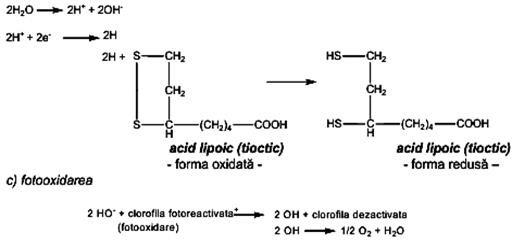
Energia necesara biosintezei poate fi realizata prin: (a) chimiosinteză, (b) fotosinteză. a) Chimiosinteza furnizează energia necesară biosintezei compuşilor organici prin oxidarea unor compuşi anorganici: H2, H2S, FeCO3, NH3, etc., cu ajutorul hidrogeno-bacteriilor, sulfobacteriilor, ferobacteriilor, bacteriilor nitrificante, etc. Importanţa biologică a chimiosintezei constă în faptul că se demonstrează posibilitatea eliberării energiei necesare sintezei materiei vii, de către celulele autotrofe, în medii minerale. b) Fotosinteza, procesul de asimilare a CO2 de către plantele verzi, este calitativ si cantitativ, cel mai important proces biochimic de pe planetă, deoarece reprezintă: • principala sursă de materie organică (se transformă C mineral din CO2 în C organic); • unica sursă de oxigen • singurul proces celular care foloseşte drept sursă energetică lumina, pe care o transformă în energie chimică. Locul de desfăşurare a fotosintezei îl reprezintă cloroplastele, produse de organite specializate ale celulelor plantelor verzi. Acestea posedă ADN propriu, care codifică partea proteică şi un tip special de membrană, denumită tilacoidă, care conţine pigmenţii clorofilieni şi sistemele transportoare de electroni. In cloroplaste, se găsesc alături de clorofile (a, b, c, d) şi pigmenţi carotenoidici, care absorb energia luminoasă şi o cedează clorofilei a, singura capabilă să o transforme în energie chimică. Moleculele clorofilelor sunt dispuse în plantele verzi sub forma unor unităţi funcţionale denumite fotosisteme, astfel încît, la absorbţia luminii să se producă rapid un transfer de energie de rezonanţă. Fotosinteza reprezintă în esenţă, un proces biologic de oxido-reducere, în care lumina este direct implicată în transferul electronilor şi în generarea de ATP. Fotosinteza se realizează în două etape greu de delimitat, datorită rapidităţii cu care se succed foarte rapid: • fotoreacţia (etapă care necesită prezenţa luminii) • scotoreacţia (etapă care nu necesită implicarea luminii la transformarea CO2 şi H2O în moleculele de glucide) Fotoreacţia, prima etapă a fotosintezei, este condiţionată de prezenţa energiei luminoase, necesară fotoactivării pigmenţilor (clorofile, carotenoide) din cloroplaste. Fotosistemele în care decurge absorbţia luminii, reacţia cu clorofila şi eliberarea electronilor, deci a poteţialului reducător, sunt implicate în două procese foarte importante: • fotoliza apei şi reducerea NADP • depozitarea energiei chimice în compusul macroergic ATP. Schematic acest proces complex, poate fi reprezentat astfel: a) fotoactivarea pigmenţilor din cloroplaste (reacţie de oxidare)
clorofila + hν (cuante de lumină) → clorofila fotoactivată+ + e-
b) fotoliza apei
c) fosforilarea fotosintetică a ADP (transformarea energiei luminoase în energie chimică)
Suma reacţiilor parţiale, se poate scrie ecuaţia globală a fotoreacţiei (ecuaţia lui Hill):
Scotoreacţia, a doua etapă a fotosintezei, poate decurge şi în absenţa luminii şi cuprinde: • reacţia de asimilare fotosintetică a CO2 absorbit prin frunze din atmosferă, sau rezultat din alte procese biochimice;
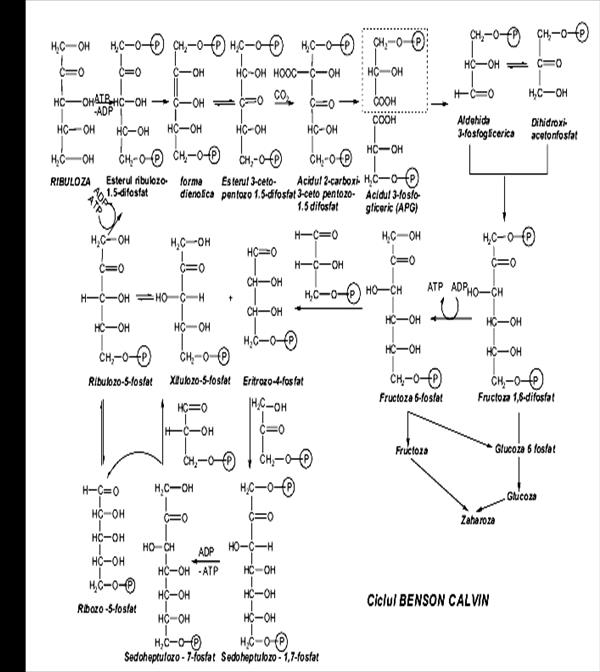
• obţinerea celor mai simple monoglucide (triozele aldehida fosfoglicerică şi hidroxoacetonfosfatul), prin participarea produselor fotoreacţiei, ATP ca activator energetic, NADPH +H+ ca agent reducător. • obţinerea primei hexoze (fructozo-1,6-difosfat) de la care se pot biosintetiza oligoglucide şi poliglucidele, sau care poate participa la regenerarea ribulozei-1,5- difosfat, necesară asimilării CO2. Succesiunea de reacţii ale scotoreacţiei constituie ciclul Benson-Calvin.
Dacă aspectul fundamental diferit al fotosintezei, faţă de alte procese biochimice, îl constituie folosirea energiei luminoase pentru a scinda fotolitic apa, a reduce NADP şi pentru a stoca energia luminoasă sub formă de energie chimică în ATP, reacţiile scotoreacţiei începînd de la metabolitul acid-3-fosfogliceric sunt asemănătoare celor produse în ficatul organismelor animale. Singura etapă caracteristică organismelor vegetale este formarea metabolitului acid 3-fosfogliceric, care implică asimilarea CO2 de către pentoza ribulozo-1,5-difosfat, prin intermediul celei mai răspîndite proteine de pe planeta-ribulozo-1,5-difosfatcarboxilaza. Şase molecule de ribulozo-1,5-difosfat (30 de atomi de C în total) şi şase molecule de CO2 (6 atomi de C) reacţionează formînd 12 molecule de acid 3-fosfogliceric (36 de atomi de C). Dintre acestea, 2 molecule (2 C3) sunt utilizate pentru a stoca glucide hexoze (C6). Cele 10 molecule de acid fosfogliceric rămase (30 atomi de C) produc 6 molecule de ribulozo-1,5-difosfat (30 atomi de C). Secvenţa de reacţii este complexă şi implică glucide pentoze, trioze, hexoze, care se transformă prin reacţii de aldolizare, de transcetolizare, etc, biocatalizate de enzime specifice.
Biosinteza pentozelor Pentozele, glucide deosebit de importante pentru organismele animale şi vegetale (riboza este componentă a acizilor nucleici şi a multor coenzime, iar ribuloza este acceptorul primar la asimilarea fotosintetică a CO2), se pot forma prin două căi metabolice: • calea (ciclul) pentozofosfaţilor (degradarea glucozo-6-fosfatului rezultat prin izomerizare din fructozo-6-fosfat) • combinarea triozelor cu aldehida acetică sau aldehida glicolică.
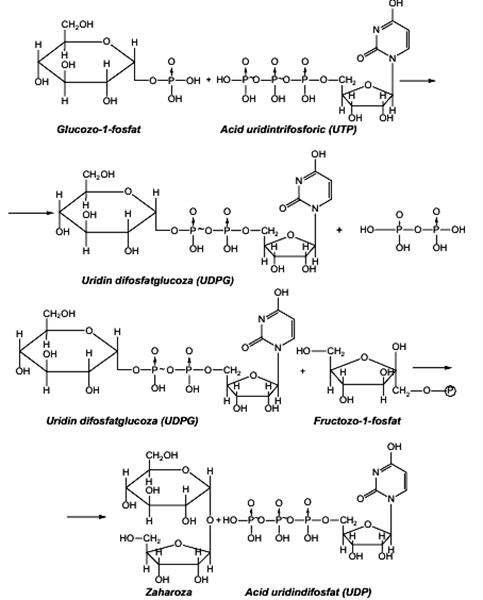
Biosinteza oligoglucidelor Biosinteza oligoglucidelor reprezintă un proces secundar al fotosintezei, respectiv transformarea hexozelor formate în diglucide, triglucide, etc. Legăturile glicozidice dintre moleculele de monoglucide pot fi realizate enzimatic prin două reacţii diferite: • transglicozilare (cazul biosintezei amilopectinei şi glicogenului) • cuplarea unor resturi de glucide activate în prealabil cu compuşi macroergici (UTP, GTP). Zaharoza (zahărul) este prima diglucidă apărută în procesul de fotosinteză şi este uşor asimilată de plante. Zaharoza se poate biosintetiza în cantităţi mici din ester glucozo-1-fosfat şi fructoză, sub influenţa biocatalitică a enzimei zaharozofosforilazei, prin eliminarea unei molecule de acid fosforic. În cantităţi mari, zaharoza se biosintetizează prin activarea uneia dintre componente (glucoza) cu compusul macroergic acid uridintrifosforic (UTP).
Diglucida maltoza se biosintetizează după un mecanism asemănător zaharozei, în care esterul glucozo-1-fosfat este transformat în uridindifosfatglucoza (UDPG) pentru a reacţiona cu glucozo-1-fosfat, cu formarea maltozei-1-fosfat.
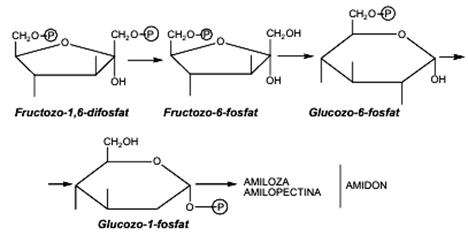
Biosinteza poliglucidelor Biosinteza poliglucidelor de rezervă glicogen (regnul animal) şi amidon (regnul vegetal) decurge cu mare viteză şi începe cu transformarea glucozo-6-fosfat în glucozo-1-fosfat, reacţie catalizată de enzima fosfoglucomutaza. Primul pas în sinteza glicogenului în organismele animale este reacţia cu compusul macroergic UTP, cu formarea uridindifosfatglucozei (UDPG), care este apoi transferată pe resturile de glucoză sau pe un fragment poliglucidic cu formarea glicogenului, sub influenţa enzimei glicogensintetaza. Glicogensintetaza necesită drept starter o catenă poliglucidică de cel puţin 4 resturi de glucoză, la care să se lege succesiv alte molecule de UDPG. Ramificarea prin legături C1 - C6 pe catena glicogenului este catalizată de o enzimă de ramificare denumită glucanaza. În plante şi în multe bacterii, sinteza amidonului decurge asemănător glicogenului, fiind catalizată de enzima amidonsintetaza.
III.1.2. Biodegradarea (catabolismul) glucidelor Biodegradarea glucidelor în organismele vii, vegetale şi animale, este un proces oxidativ, generator de energie (exergonic), care constă în transformarea moleculelor de glucide (poliglucide, oligoglucide, monoglucide) în compuşi mai simpli, cu masă moleculară mai mică. Biodegradarea glucidelor poate decurge, în funcţie de condiţii, în două etape: • biodegradarea (catabolismul) anaerobă • biodegradarea (catabolismul) aerobă Catabolismul anaerob decurge în organismele vii în absenţa oxigenului, pe două căi metabolice: • biodegradarea glicolitică (glicoliza), un ciclu de reacţii prin care glucoza este transformată în acid piruvic şi se formează ATP. • Biodegradări fermentative: (a) fermentaţia anaerobă de diferite tipuri (alcoolică, lactică, butirică, etc.) avînd ca produs final: alcoolul etilic, acidul lactic, acidul butiric şi CO2, respectiv, fermentaţia aerobă. Biodegradarea glicolitică a glucidelor (glicoliza) una dintre căile metabolice care demonstrează universitalitatea proceselor biochimice, a fost complet elucidată pînă în anul 1940 (mai ales datorită savanţilor G. Embden, O. Meyerhoff, J, Parnas, de unde şi denumirea ciclului de transformări ciclul Embden - Meyerhoff - Parnas. Toate reacţiile glicolizei decurg în citosolul celulelor. Procesul de biodegradare a poli-glucidei amidon decurge prin parcurgerea următoarelor etape: • scindarea legăturilor C1 –C6 glicozidice din amiloză pe cale hidrolitică (sub acţiunea enzimei amilaza), sau pe cale fosforolitică (sub acţiunea enzimei transglucozidaza), cu eliberarea unei molecule de ester glucozo-1-fosfat; • legăturile C1–C6 glicozidice din amilopectina sunt scindate hidrolitic de către enzima amilo-1,6-glucozidaza; • în cazul în care glicoliza porneşte direct de la glucoză este necesară fosforilarea prealabilă cu ATP în prezenţa enzimei hexokinaza (activată de prezenţa ionilor de Mg2+ sau Mn2+) cu formarea esterului glucozo-1-fosfat; • esterul glucozo-1-fosfat (ester Cori) este izomerizat la ester glucozo-6-fosfat (ester Robinson) sub acţiunea fosfoglucomutazei; • următoarea etapă a glicolizei constă în izomerizarea glucozo-6-fosfatului la fructozo-6-fosfat (ester Neuberg). Aceasta este o reacţie de conversie a unei aldoze într-o cetoză, sub acţiunea enzimei fosfohexozizomeraza; • are loc a doua reacţie de fosforilare (sub acţiunea ATP şi a fosfofructokinazei) a fructozo-6-fosfatului cu formarea fructozo-1,6-difosfatului (ester Harden-Young), ultima glucidă cu şase atomi de carbon din ciclul glicolizei; • sub acţiunea biocatalitică a enzimei aldolaza, esterul fructozo-1,6-difosfat este scindat în aldehida 3-fosfoglicerica şi dihidroxiacetonfosfat. Toate celelalte etape ale glicolizei decurg numai cu unităţi formate din cîte 3 atomi de carbon; • glicoliza decurge în continuare prin intermediul aldehidei-3-fosfoglicerice, la care este convertit şi dihidroxiacetonfosfatul, sub acţiunea fosfotriozizomerazei.In etapele precedente, glicoliza a decurs cu scindarea unei molecule de glucoză în două molecule de aldehidă 3-fosfoglicerică, cu consumarea a doi moli de ATP: • urmează o etapă foarte importantă din punct de vedere al producerii de energie- fosforilarea oxidativă a aldehidei-3-fosfoglicerice în acid 1,3-difosfogliceric, care decurge în prezenţa enzimei fosfogliceroaldehiddehidrogenazei avînd drept coenzimă NAD. Energia eliberată în această reacţie de oxido-reducere se înmagazinează în ATP prin legătura macroergică apărută între ADP si H3PO4; • în ultima etapă a glicolizei se formează acidul piruvic şi o a doua moleculă de ATP printr-o secvenţă de reacţii care implică: - o reacţie de defosforilare (în prezenţa ADP, a fosfoglicerokinazei şi a ionului Mg2+) a acidului 1,3-difosfogliceric la acid 3-fosfogliceric (ester Nilson).
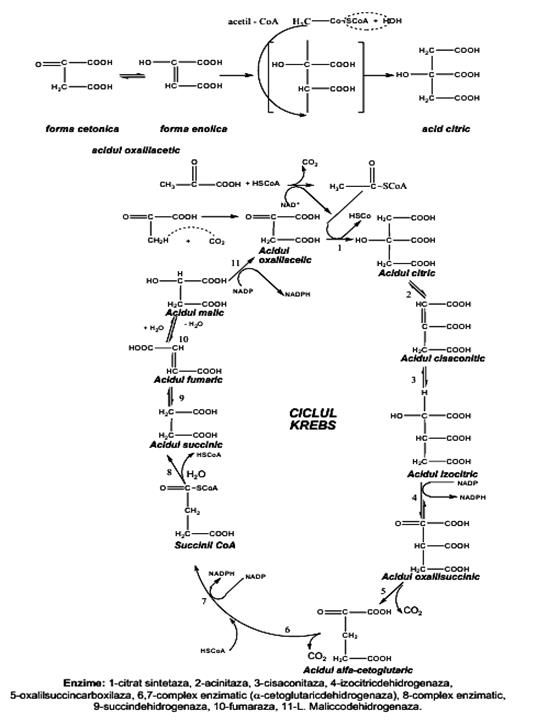
- izomerizarea acidului 3-fosfogliceric la acid 2-fosfogliceric, reacţie catalizată de enzima fosfogliceromutaza. - transformarea acidului 2-fosfogliceric în acid 2-fosfoenolpiruvic, sub acţiunea enolazei. - defosforilarea acidului 2-fosfoenolpiruvic sub acţiunea piruvatkinazei, în prezenţă de ADP, Mg2+ şi K+, cu formarea acidului piruvic şi a ATP. Acidul piruvic este unul dintre cei mai importanţi metaboliţi, finalul biodegradării anaerobe şi compusul de la care începe biodegradarea aerobă a glucidelor. El reprezintă, de asemenea, o punte de legătura cu protidele şi cu lipidele. Glicoliza este reprezentată în schema de reacţii următoare prezentat schema 3.1 Biodegradarea aerobă a glucidelor reprezintă etapa de desăvîrşire a biodegradării glucidelor şi decurge prin transformarea acidului piruvic, produsul final al glicolizei, în acetilcoenzima A, care va fi, în final, oxidată total la CO2 şi H2O. Ciclul acidului citric (sau ciclul Krebs, sau ciclul acizilor tricarboxilici) reprezintă calea metabolică comună de descompunere oxidativă a glucidelor, lipidelor şi aminoacizilor. Principiul de funcţionare a metabolismului poate fi enunţat astfel: • moleculele organice complicate sunt scindate în unităţi C2 (acid acetic activat) • biodegradarea unităţilor C2 conduce la formarea a doi moli de CO2 şi 2 atomi de H.
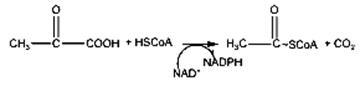
Schema 3.1 Glicoliza • produsul final H2O apare în lanţul respirator din atomii de hidrogen de pe coenzime şi oxigen, cu depozitarea unei părţi din energie sub forma compusului ATP. • spre deosebire de glicoliză, care decurge în citosol, reacţiile ciclului Krebs decurg în mitocondrii - • biodegradarea aerobă a glucidelor este precedată de 2 reacţii importante: - decarboxilarea acidului piruvic cu formarea acetilcoenzimei A, compus macroergic - carboxilarea acidului piruvic la acid oxalilacetic, care prin reacţia cu acetilcoenzima A va forma acidul citric. Decarboxilarea acidului piruvic reprezintă etapa de legătura între glicoliză şi ciclul acidului citric. Reacţia este catalizată de enzima piruvatdehidrogenaza şi poate fi reprezentată astfel:
Decarboxilarea se produce printr-o succesiune de reacţii intermediare, catalizate fiecare de cîte o enzimă din sistemul multienzimatic al piruvatdehidrogenazei, cu participarea coenzimelor specifice: tiaminpirofosfatul (TPP), NADP+, HSCoA, acidul lipoic, Mg2+. Prima etapă constă în decarboxilarea acidului piruvic, cu producerea CO2 şi a unei grupe hidroxietil ataşată tiaminpirofosfatului. Grupa hidroxietil este transformată în acetilcoenzima A, cu reducerea NAD+, proces denumit decarboxilare oxidativă. Conversia acidului piruvic la acetilcoenzima A este ireversibilă, ca urmare acizii graşi superiori nu pot fi transformaţi în glucoză în organismele animale, deşi bacteriile şi plantele o pot face, printr-un mecanism special. Atomii de H fixaţi de NADP+ si FAD în etapa de decarboxilare oxidativă şi rezultaţi şi din alte etape ale ciclului, se vor combina cu oxigenul, cu formarea apei (în aşa numitul ciclu respirator). Carboxilarea acidului piruvic este o etapă în care are loc fixarea CO2 activat cu biotinilenzima şi cu ATP la acidul piruvic, cu formarea acidului oxalilacetic.
Biosinteza acidului citric decurge ireversibil, prin condensarea aldolică a acetilcoenzimei A cu acidul oxalilacetic, sub influenţa enzimei citratsintetaza. Energia necesară reacţiei este furnizată prin hidroliza legăturii macroergice din acetilcoenzima A. • deshidratarea acidului citric la acid cis-aconitic şi hidratarea la acid izocitric (ambele etape) sunt catalizate de enzima aconitaza. • urmează prima din cele patru reacţii de oxido-reducere din ciclul Krebs şi anume, dehidrogenarea acidului izocitric cu ajutorul enzimei izocitricdehidrogenaza, avînd drept coenzima NADP+ şi formarea acidului oxalilsuccinic.
• prima decarboxilare din ciclu, decurge la acidul oxalilsuccinic, un Я-acid instabil, care se decarboxilează cu ajutorul izocitricdehidrogenazei, cu formarea acidului cetoglutaric. • a doua decarboxilare oxidativă din ciclu, a acidului cetoglutaric la succinilcoenzima A, care prin hidroliză formează acidul succinic, decurge cu participarea aceluiaşi sistem enzimatic întîlnit la etapa de decarboxilare a acidului piruvic la acetilcoenzima A (α-cetoglutaricdehidrogenaza) • ultimele reacţii ale ciclului constau în transformarea în trei etape a acidului succinic în acid oxalilacetic: - oxidarea (dehidrogenarea cu FAD) a acidului succinic la acid fumaric - hidratarea acidului fumaric la acid malic - oxidarea (dehidrogenarea cu NAD+) a acidului malic la acid oxalilacetic Acidul oxalilacetic regenerat poate asigura continuarea aceleiaşi succesiuni de reacţii, cu eliberare de energie stocata sub forma de FADH şi NAD+. Bilantul general al ciclului Krebs poate fi reprezentat astfel:
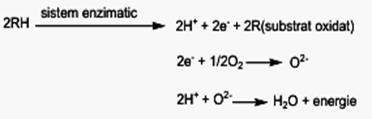
Sensul metabolic al ciclului constă în: • oxidarea restului acetil (CH3CO-) din acetilcoenzima A rezultată din acid piruvic, la CO2 , substanţă оn care carbonul este complet oxidat la C4+. • din cele patru reacţii de oxidare rezultă 8 atomi de H, legaţi de coenzimele NAD+ şi FAD, care sunt participanţi la lanţul respirator, unde de-a lungul unui sistem enzimatic specific, suferă reacţii de oxidare la H+, formă sub care reacţionează cu O2-, formînd H2O şi eliberînd energie. Procesul de respiraţie tisulară (lanţul respirator) poate fi reprezentatat, sumar, astfel:
Ciclul Krebs(Schema 3.2) operează în sens unic datorită celor trei reacţii care decurg ireversibil: • sinteza acidului citric din acetilcoenzima A şi acid oxalilacetic • decarboxilarea acidului izocitric la acid α-cetoglutaric • dehidrogenarea acidului α-cetoglutaric Energia eliberată treptat, în cursul reacţiilor este înmagazinată sub forma legăturilor macroergice ale compuşilor macroergici GTP, ATP şi eliberată treptat în funcţie de nevoile organismului (procese de biosinteză, energie osmotică, electrică, mecanică, calorică, etc.). În următoarea schemă este reprezentată succesiunea reacţiilor din ciclul Krebs. Importanţa ciclului Krebs decurge din faptul că, constituie o cale comună de biodegradare a glucidelor, a lipidelor si protidelor (prin substanţele intermediare provenite din acestea). Cetoacizii rezultaţi în ciclul citric, pot servi la biosinteza aminoacizilor (corelaţie între glucide şi protide), iar prin intermediul acetilcoenzimei A se stabileşte corelaţia metabolică dintre glucide şi lipide. Se poate concluziona că, prin interrelaţiile metabolice stabilite între glucide-lipide-protide, ciclul Krebs reprezintă o sursă de substrat şi de energie necesară tuturor proceselor metabolice. Biodegradarea fermentativă a glucidelor Biodegradările fermentative sunt procese de descompunere, predominant anaerobă, ale glucidelor datorate activităţii fiziologice a unor microorganisme. Au o importanţă practică deosebită, contribuind la circuitul elementelor în natură, la fertilizarea solului, la obţinerea unor compuşi utilizaţi în industria alimentară, chimică, farmaceutică.
Fermentaţiile anaerobe (în absenţa oxigenului) Fermentaţiile anaerobe (în absenţa oxigenului) se pot clasifica, după natura substanţelor finale în: - fermentaţia alcoolică - fermentaţia lactică - fermentaţia butirică - fermentaţia propionică
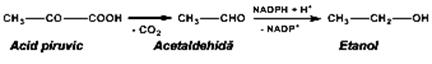
Schema 3.2 Chiclul Krebs a ) Fermentaţia alcoolică a glucidelor decurge sub influenţa drojdiilor Saccharomices cerevisiae. Mecanismul este identic cu cel al glicolizei, pînă la acidul piruvic. În continuare, acidul piruvic este decarboxilat la acetaldehidă, (în prezenţa catalizatorului piruvatdecarboxilaza, avînd drept coenzimă tiaminpirofosfatul. Acetaldehida este apoi redusă la etanol cu enzima alcooldehidrogenaza şi NADH + H+.
În decursul fermentaţiei alcoolice rezultă şi produşi secundari: acetaldehidă, acid lactic, alcool amilic, acid succinic, etc., care trebuiesc separaţi prin distilare fracţionată.
b ) Fermentaţia lactică este întîlnită în mod normal în diferite microorganisme, dar şi în muşchi la efort intens depus în condiţii de concentraţii de reduse de oxigen. Mecanismul fermentaţiei lactice este analog glicolizei, pînă la acidul piruvic. Mai departe, acidul piruvic se transformă printr-un proces de reducere enzimatică în acid lactic sub acţiunea enzimei lactatdehidrogenaza avînd drept coenzima NADH+H+.
Fermentaţia lactică se aplică în industria alimentară la prepararea iaurtului, murăturilor, panificaţie, etc.). c ) Fermentaţia propionică este un proces de degradare anaerobă a glucidelor, care decurge sub acţiunea unor bacterii specifice, în urma căruia se obţine, comparativ cu fermentaţia alcoolică şi cea lactică, o cantitate mai mare de ATP, respectiv, cîte 2 moli de ATP/mol de glucoză.
Fermentaţia propionică este utilizată, alături de fermentaţia lactică, în industria alimentară la fabricarea brînzeturilor, pentru activarea unor procese biochimice din sol, etc. d) Fermentaţia butirică constă în bioegradarea glucidelor de către anumite microorganisme, cu formarea acidului butiric.
Decurge cu descompunerea unei mari cantităţi de substanţă organă, în soluri mlăştinoase, în nămoluri, în bălţi, cu degajare de miros neplăcut.
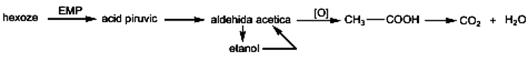
Fermentaţia aerobă (în prezenţa oxigenului) Fermentaţia aerobă este un proces biochimic care decurge în prezenţa oxigenului. Mecanismul de reacţie este similar cu cel al fermentaţiei alcoolice, pînă la etapa de formare a aldehidei acetice, după care, în urma procesului de oxidare, se formează acid acetic. Acidul acetic poate fi biodegradat pînă la CO2 şi H2O.
|
|||||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 451; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.118.148.168 (0.097 с.) |