Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Понятие о тканях. Классификация
Ткани – этоустойчивые, т.е. закономерно повторяющиеся комплексы клеток, сходные по происхождению, строению и приспособленные к выполнению одной или нескольких функций. Ткани возникли у растений в связи с переходом их предков – водорослей к наземному образу жизни. Переход от условий водной среды к условиям суши сопровождался дифференциацией однородного слоевища водных обитателей на основные вегетативные органы: корень и побег. Возникли группы клеток, выполняющие определенные функции. В процессе эволюции строение тела растений усложнялось: у мхов имеется около 20 различных типов клеток, у папоротникообразных – около 40, у покрытосеменных – более 80. Наиболее совершенные и сложные по структуре ткани сформировались у цветковых растений. Ткани классифицируют в зависимости от выполняемой функции. Однако надо помнить, что одна и та же ткань может выполнять несколько функций. Различают сложные ткани, состоящие из разнородных элементов, выполняющих разные функции, и простые ткани, состоящие из однородных элементов. Функции ткани могут меняться в зависимости от ее возраста. Обычно ткани локализуются в теле растения определенным образом, образуя единые массивы. Иногда клетки, относящиеся к одной ткани, могут быть рассеяны поодиночке среди клеток других тканей и разобщены между собой. Такие клетки называются идиобластами (например секреторные клетки, склереиды). Классификации тканей довольно разнообразны. Наиболее часто выделяют шесть типов тканей: 1) образовательные, или меристемы; 2) основные; 3) покровные; 4) выделительные; 5) механические; 6) проводящие. Последние пять типов объединяют в постоянные ткани. Они образуются из меристем путем дифференциации клеток.
Образовательные ткани, или меристемы Тело растения формируется в результате деятельности образовательных тканей, называемых меристемами. Основное свойство меристем – способность к делению и образованию новых клеток. В теле растения образовательные ткани функционируют в течение всей жизни. У векового дерева, наряду с очень старыми тканями можно обнаружить и молодые. В составе меристем различают: 1) инициальные клетки, или инициали, и 2) производные от инициалей. Инициали сохраняют способность к делению в течение всей жизни растения и всегда остаются в составе меристем. Производные от инициалей делятся некоторое число раз и затем превращаются в постоянные ткани.
По происхождению различают: 1) первичные меристемы, которые берут начало непосредственно от меристем зародыша, и 2) вторичные меристемы, образующиеся на более поздних этапах развития растения либо из первичных меристем, либо в результате дедифференциации постоянных тканей. Постоянные ткани, образовавшиеся из первичных меристем, называются первичными, из них складывается первичная структура тела растения. Из вторичных меристем образуются вторичные ткани, которые определяют вторичный рост растения. В зависимости от местоположения выделяют четыре типа меристем: 1) верхушечные, или апикальные; 2) боковые, или латеральные; 3) вставочные, или интеркалярные; 4) раневые, или травматические. Верхушечные (апикальные) меристемы закладываются с первых стадий развития зародыша на верхушке побега и на кончике зародышевого корешка. По мере роста и ветвления на каждом боковом побеге и каждом корне образуются свои верхушечные (апикальные) меристемы. Они обеспечивают рост этих органов в длину. Апикальные меристемы всегда первичны, они образуют конусы нарастания корня и побега. Боковые (латеральные) меристемы располагаются по окружности осевых органов (корней, стеблей) в виде цилиндров, которые на поперечных срезах имеют вид колец. Первичные боковые меристемы – прокамбий, перицикл – возникают непосредственно под апексами и в непосредственной связи с ними. Вторичные латеральные меристемы: камбий – возникает из прокамбия - и пробковый камбий (феллоген) - образуется из клеток постоянных тканей. Боковые меристемы обеспечивают рост корня и стебля в толщину. Из прокамбия и камбия образуются проводящие ткани, из феллогена – перидерма. Вставочные (интеркалярные) меристемы находятся в основаниях междоузлий побегов и молодых листьев. Они первичны, поскольку являются остатками верхушечных меристем, их дифференциация задерживается по сравнению с остальными тканями. Вставочные меристемы не имеют в своем составе инициалей и со временем полностью превращаются в постоянные ткани. В их составе могут присутствовать некоторые дифференцированные элементы, например проводящие. Интеркалярный рост характерен для стеблей злаков, наблюдается также в основании луковиц, завязей.
Раневые (травматические) меристемы обычно образуются при повреждении тканей и органов. Живые клетки постоянных тканей, окружающие пораженные участки, дедифференцируются и начинают делиться, т.е. превращаются во вторичную меристему. Раневые меристемы образуют каллус –особую ткань, состоящую из однородных паренхимных клеток, прикрывающих место поранения. Из каллуса может возникнуть любая ткань или орган растения. Часто клетки формируют феллоген, образующий на поверхности перидерму, которая закрывает рану и способствует ее заживлению. Способность растений к каллусообразованию используют для получения культуры изолированных тканей, а также в практике садоводства для размножения растений черенками и прививками. Типичные цитологические признаки образовательных тканей наиболее отчетливо выражены у апикальных меристем. Это изодиаметрические многогранные клетки, не разделенные межклетниками. Клеточные стенки тонкие, с малым содержанием целлюлозы. Цитоплазма густая, ядро относительно крупное, занимает центральное положение. В цитоплазме большое число рибосом и митохондрий, так как происходит интенсивный синтез белков и других веществ. Вакуоли очень мелкие, многочисленные (рис. 3.1).
Рис. 3.1. Верхушечная меристема побега элодеи: А – продольный разрез; Б – внешний вид и продольный разрез конуса нарастания; В – клетки первичной меристемы; Г – паренхимная клетка листа, закончившая дифференцировку; 1 – конус нарастания; 2 – зачаток листа; 3 – зачаток бокового побега.
Основные ткани Основные ткани составляют основную массу тела растения. Они состоят из живых, относительно мало специализированных клеток, чаще паренхимной формы, поэтому их часто называют паренхимными тканями, или паренхимой. В зависимости от выполняемой функции, различают несколько типов основных тканей. Ассимиляционная ткань (хлорофиллоносная паренхима, хлоренхима) выполняет функцию фотосинтеза. Она располагается в основном в листьях и стеблях травянистых растений сразу за эпидермой. Клетки живые, тонкостенные, чаще паренхимной формы. 70-80% объема протопласта составляют хлоропласты. Характерно наличие межклетников, которые облегчают газообмен (рис. 3.2).
Рис. 3.2. Поперечный срез листа красавки: 1 – клетки ассимиляционной ткани; 2 – клетки, заполненные кристаллическим песком кальция оксалата.
Запасающая паренхима служит местом отложения питательных веществ (крахмала, белков, жирных масел). Запасные питательные вещества могут откладываться в живых клетках любой ткани, но особенно ярко эта функция проявляется у специализированных запасающих тканей, хорошо развитых в семенах, корнях, подземных побегах (рис. 3.3.А). Состоят запасающие ткани из живых тонкостенных клеток, чаще паренхимной формы. Разновидностью запасающей ткани является водоносная паренхима, выполняющая функцию запасания воды. Она состоит из крупных живых тонкостенных клеток, как правило, паренхимной формы. Вода запасается в вакуолях за счет большого содержания слизей, обладающих высокой водоудерживающей способностью. Водоносная паренхима имеется в стеблях и листьях суккулентов (кактусы, агавы, алоэ), у многих растений солончаков (солерос, анабазис, саксаул), в листьях многих злаков. Много воды содержится в запасающих тканях луковиц и клубней.
Воздухоносная паренхима (аэренхима) выполняет функцию вентиляции, снабжая ткани и органы кислородом. Она хорошо развита в погруженных органах водных и болотных растений (кувшинка, кубышка, аир, вахта). Аэренхима состоит из живых клеток различной формы и крупных межклетников (рис. 3.3.Б).
Рис. 3.3. Запасающая паренхима клубня картофеля (A) и аэренхима стебля рдеста (Б): 1 – межклетник.
Механическая паренхима занимает промежуточное положение между основными и механическими тканями. Это живые паренхимные клетки со слегка утолщенной одревесневшей клеточной стенкой. Неспециализированная паренхима (основная паренхима, неспецифическая паренхима) представляет собой живую паренхимную ткань без выраженной функции. Эта ткань всегда присутствует в теле растения, составляя его большую часть.
Покровные ткани Покровные ткани располагаются на поверхности органов растений на границе с внешней средой. Они состоят из плотно сомкнутых клеток и защищают внутренние части растения от неблагоприятных внешних воздействий, излишнего испарения и иссушения, резкой перемены температуры, проникновения микроорганизмов, служат для газообмена и транспирации. В соответствии с происхождением из различных меристем выделяют первичные и вторичные покровные ткани. К первичным покровным тканям относят: 1) ризодерму, или эпиблему и 2) эпидерму. Ризодерма (эпиблема) – первичная однослойная поверхностная ткань корня. Образуется из протодермы – наружного слоя клеток апикальной меристемы корня. Основная функция ризодермы – всасывание, избирательное поглощение из почвы воды с растворенными в ней элементами минерального питания. Через ризодерму происходит выделение веществ, действующих на субстрат и преобразующих его. Клетки ризодермы тонкостенные, с вязкой цитоплазмой и большим количеством митохондрий (минеральные ионы поглощаются активно, с затратой энергии, против градиента концентрации). Характерной особенностью ризодермы является образование у части клеток корневых волосков – трубчатых выростов, в отличие от трихомов не отделенных стенкой от материнской клетки (рис. 3.4). Корневые волоски увеличивают поглощающую поверхность ризодермы в десять и более раз. Волоски имеют длину 1-2 (3) мм. Ризодерму часто рассматривают как всасывающую ткань.
Рис. 3.4. Кончик корня ожики многоцветковой: 1 – корневой волосок.
Эпидерма - первичная покровная ткань, образующаяся из протодермы конуса нарастания побега. Она покрывает листья, стебли травянистых и молодых побегов древесных растений, цветки, плоды и семена. Основная функция эпидермы – регуляция газообмена и транспирации (испарения воды живыми тканями). Кроме того, эпидерма выполняет целый ряд других функций. Она препятствует проникновению внутрь растения болезнетворных организмов, защищает внутренние ткани от механических повреждений и придает органам прочность. Через эпидерму могут выделяться наружу эфирные масла, вода, соли. Эпидерма может функционировать как всасывающая ткань. Она принимает участие в синтезе различных веществ, в восприятии раздражений, в движении листьев. Эпидерма - сложная ткань, в ее состав входят морфологически различные типы клеток: 1) основные клетки эпидермы; 2) замыкающие и побочные клетки устьиц; 3) трихомы. Основные клетки эпидермы – живые клетки таблитчатой формы. Вид клеток с поверхности различен (рис. 3.5). Клетки плотно сомкнуты, межклетники отсутствуют. Боковые стенки (перпендикулярные поверхности органа) часто извилистые, что повышает прочность их сцепления, реже прямые. Эпидермальные клетки осевых органов и листьев многих однодольных сильно вытянуты вдоль оси органа. Рис. 3.5. Эпидерма листа различных растений (вид с поверхности): 1 - ирис; 2 - кукуруза; 3 – арбуз; 4 - буквица.
Наружные стенки клеток обычно толще остальных. Их внутренний, более мощный, слой состоит из целлюлозы и пектиновых веществ; наружный слой подвергается кутинизации. Поверх наружных стенок выделяется сплошной слой кутина, образующий защитную пленку – кутикулу. Помимо кутина в ее состав входят вкрапления воска, что еще больше снижает проницаемость кутикулы для воды и для газов. Воск может откладываться в кристаллической форме и на поверхности кутикулы в виде чешуек, палочек, трубочек и других структур, видимых только в электронный микроскоп. Этот сизый, легко стирающийся налет хорошо заметен на листьях капусты, плодах сливы, винограда. Мощность кутикулы, распределение в ней восков и кутина определяют химическую стойкость и проницаемость эпидермы для газов и растворов. В условиях засушливого климата у растений развивается более толстая кутикула. У растений, погруженных в воду, кутикула отсутствует. Клетки эпидермы имеют живой протопласт, обычно с хорошо развитой эндоплазматической сетью и аппаратом Гольджи. У большинства видов растений в цитоплазме присутствуют лейкопласты. У водных растений, папоротников, обитателей тенистых мест (гибискус) встречаются редкие хлоропласты. Эпидерма чаще всего состоит из одного слоя клеток. Редко встречается двух- или многослойная эпидерма, преимущественно у тропических растений, живущих в условиях непостоянной обеспеченности водой (бегонии, пеперомии, фикусы). Нижние слои многослойной эпидермы функционируют как водозапасающая ткань. У некоторых растений клеточные стенки могут пропитываться кремнеземом (хвощи, злаки, осоки) или содержать слизи (семена льна, айвы, подорожников).
Устьица – образования для регуляции транспирации и газообмена. Устьице состоит из двух замыкающих клеток бобовидной формы, между которыми находится устьичная щель, которая может расширяться и сужаться. Под щелью располагается крупный межклетник – подустьичная полость. Клетки эпидермы, примыкающие к замыкающим клеткам, часто отличаются от остальных клеток, и тогда их называют побочными, или околоустьичными клетками (рис. 3.6). Они участвуют в движении замыкающих клеток.
Рис. 3.6. Схема строения устьица.
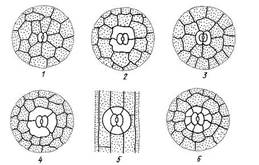
Замыкающие и побочные клетки образуют устьичный аппарат. В зависимости от числа побочных клеток и их расположения относительно устьичной щели выделяют несколько типов устьичного аппарата (рис. 3.7). В фармакогнозии типы устьичного аппарата используются для диагностики лекарственного растительного сырья.
Рис. 3.7. Типы устьичного аппарата: 1 – аномоцитный; 2 – диацитный; 3 – парацитный; 4 – анизоцитный; 5 – тетрацитный; 5 – энциклоцитный.
Аномоцитный тип устьичного аппарата обычен для всех групп растений, исключая хвощи. Побочные клетки в этом случае не отличаются от остальных клеток эпидермы. Диацитный тип характеризуется двумя побочными клетками, которые располагаются перпендикулярно устьичной щели. Этот тип обнаружен у некоторых цветковых растений, в частности, у большинства губоцветных (мята, шалфей, чабрец, душица) и гвоздичных. При парацитном типе две побочные клетки располагаются параллельно замыкающим и устьичной щели. Он найден у папоротников, хвощей и ряда цветковых растений. Анизоцитный тип обнаружен только у цветковых растений, в частности, он встречается у крестоцветных (пастушья сумка, желтушник) и пасленовых (белена, дурман, красавка). В этом случае замыкающие клетки окружены тремя побочными, одна из которых заметно крупнее или мельче остальных. Тетрацитным типом устьичного аппарата характеризуются преимущественно однодольные. При энциклоцитном типе побочные клетки образуют узкое кольцо вокруг замыкающих клеток. Подобная структура найдена у папоротников, голосеменных и некоторых цветковых. Механизм движения замыкающих клеток основан на том, что стенки их утолщены неравномерно, поэтому форма клеток меняется при изменении их объема. Изменение объема клеток устьичного аппарата происходит вследствие изменения осмотического давления. Увеличение давления происходит за счет активного поступления из соседних клеток ионов калия, а также за счет повышения концентрации сахаров, образующихся в процессе фотосинтеза. За счет поступления воды объем вакуоли увеличивается, тургорное давление растет, и устьичная щель открывается. Отток ионов совершается пассивно, вода выходит из замыкающих клеток, их объем уменьшается, и устьичная щель закрывается. У большинства растений устьица открываются в светлое время суток и закрываются ночью. Это связано с тем, что фотосинтез протекает только на свету, и для него необходим приток из атмосферы углекислого газа. Число и распределение устьиц очень варьируют в зависимости от вида растения и экологических условий. У большинства растений их число составляет 100-700 на 1мм2 поверхности листа. С помощью устьиц эпидерма эффективно регулирует газообмен и транспирацию. Если устьица полностью открыты, то транспирация идет с такой же скоростью, как если бы эпидермы не было вовсе (согласно закону Дальтона, при одной и той же суммарной площади отверстий скорость испарения тем выше, чем больше число отверстий). При закрытых устьицах транспирация резко снижается и фактически может идти только через кутикулу. У многих растений эпидерма образует наружные одно- или многоклеточные выросты различной формы – трихомы. Трихомы отличаются крайним разнообразием, оставаясь вместе с тем вполне устойчивыми и типичными для определенных видов, родов и даже семейств. Поэтому признаки трихомов широко используются в систематике растений и в фармакогнозии в качестве диагностических. Трихомы делятся на: 1) кроющие и 2) железистые. Железистые трихомы образуют вещества, которые рассматриваются как выделения. Они будут рассмотрены в разделе, посвященном выделительным тканям. Кроющие трихомы имеют вид простых, разветвленных или звездчатых волосков, одно- или многоклеточных (рис. 3.8). Кроющие трихомы могут длительное время оставаться живыми, но чаще они быстро отмирают и заполняются воздухом. Густой слой волосков отражает часть солнечных лучей и уменьшает нагрев, создает затишное пространство около эпидермы, что в совокупности снижает транспирацию. Часто волоски образуют покров только там, где располагаются устьица, например на нижней стороне листьев мать-и-мачехи, багульника. Жесткие, колючие волоски защищают растения от поедания животными, сосочки на лепестках привлекают насекомых.
Рис. 3.8. Кроющие трихомы: 1-3 – простые одноклеточные, 4 – простой многоклеточный, 5 – ветвистый многоклеточный, 6 – простой двурогий, 7,8 – звездчатый (в плане и на поперечном разрезе листа).
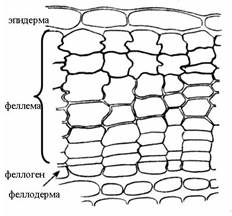
От трихомов, образующихся только из эпидермальных клеток, следует отличать эмергенцы, в формировании которых принимают участие и более глубоко расположенные ткани. К ним относят шипы розы, малины, ежевики, покрывающие черешки листьев и молодые побеги. К вторичным покровным тканям относятся: 1) перидерма и 2) корка, или ритидом. Перидерма – сложная многослойная покровная ткань, которая приходит на смену первичным покровным тканям – ризодерме и эпидерме. Перидерма покрывает корни вторичного строения и стебли многолетних побегов. Она может возникнуть и в результате залечивания поврежденных тканей раневой меристемой. Перидерма состоит из трех комплексов клеток, различных по строению и функциям. Это: 1) феллема, или пробка, выполняющая главные защитные функции; 2) феллоген, или пробковый камбий, за счет работы которого образуется перидерма в целом; 3) феллодерма, или пробковая паренхима, выполняющая функцию питания феллогена (рис. 3.9).
Рис. 3.9. Строение перидермы стебля бузины.
Феллема (пробка) состоит из нескольких слоев таблитчатых клеток, расположенных плотно, без межклетников. Вторичные клеточные стенки состоят из чередующихся слоев суберина и воска, что делает их непроницаемыми для воды и газов. Клетки пробки мертвые, они не имеют протопласта и заполнены воздухом. В полости клеток могут также откладываться вещества, повышающие защитные свойства пробки. Феллоген (пробковый камбий) – вторичная латеральная меристема. Это один слой меристематических клеток, откладывающих клетки пробки наружу и клетки феллодермы внутрь органа. Феллодерма (пробковая паренхима) относится к основным тканям и состоит из живых паренхимных клеток. Однако часто феллоген работает односторонне, откладывая только пробку, а феллодерма остается однослойной (рис. 3.9). Главная функция пробки – защита от потери влаги. Кроме того, пробка предохраняет растение от проникновения болезнетворных организмов, а также дает механическую защиту стволам и ветвям деревьев, а феллоген залечивает нанесенные повреждения, образуя новые слои пробки. Поскольку клетки пробки заполнены воздухом, пробковый футляр обладает малой теплопроводностью и хорошо предохраняет от резких колебаний температуры. У большинства деревьев и кустарников феллоген закладывается в однолетних побегах уже в середине лета. Чаще всего он возникает из паренхимных клеток, лежащих сразу под эпидермой (рис. 3.9). Иногда феллоген образуется в более глубоких слоях коры (смородина, малина). Редко эпидермальные клетки, делясь, превращаются в феллоген (ива, айва, олеандр). Газообмен и транспирация в органах, покрытых перидермой, происходят через чечевички (рис. 3.10). В местах чечевичек пробковые слои разорваны и чередуются с паренхимными клетками, рыхло соединенными между собой. По межклетникам этой выполняющей ткани циркулируют газы. Феллоген подстилает выполняющую ткань и, по мере ее отмирания, дополняет новыми слоями. С наступлением холодного сезона феллоген откладывает под выполняющей тканью замыкающий слой, состоящий из клеток пробки. Весной этот слой под напором новых клеток разрывается. В замыкающих слоях имеются небольшие межклетники, так что живые ткани ветвей деревьев даже зимой не отграничены наглухо от окружающей среды.
Рис. 3.10. Строение чечевички бузины на поперечном разрезе.
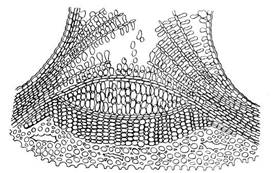
На молодых побегах чечевички выглядят как небольшие бугорки. По мере утолщения ветвей их форма меняется. У березы они растягиваются по окружности ствола и образуют характерный рисунок из черных черточек на белом фоне. У осины чечевички принимают форму ромбов. У большинства древесных растений на смену гладкой перидерме приходит трещиноватая корка (ритидом). У сосны это происходит на 8-10-м году, у дуба – в 25-30 лет, у граба – в 50 лет. Лишь у некоторых деревьев (осина, бук, платан, эвкалипт) корка вообще не образуется. Корка возникает в результате многократного заложения новых прослоек перидермы во все более глубоких слоях коры. Живые клетки, заключенные между этими прослойками, погибают. Таким образом, корка состоит из чередующихся слоев пробки и прочих отмерших тканей коры (рис. 3.11).
Рис. 3.11. Корка дуба на поперечном разрезе.
Мертвые ткани корки не могут растягиваться, следуя за утолщением ствола, поэтому на стволе появляются трещины, не доходящие, однако, до глубинных живых тканей. Граница между перидермой и коркой внешне заметна по появлению этих трещин, особенно ясна эта граница у березы, у которой белая береста (перидерма) сменяется черной трещиноватой коркой. Толстая корка надежно предохраняет стволы деревьев от механических повреждений, лесных пожаров, резкой смены температур.
Механические ткани Механические (скелетные, опорные, арматурные) ткани выполняют в растении роль скелета, который скрепляет ткани и части органов между собой. Они придают растениям прочность, способность противостоять действию тяжести собственных органов, порывам ветра, дождю, снегу, вытаптыванию животными. Клетки механических тканей разнообразны по форме, но имеют общий признак – сильно утолщенные клеточные стенки, которые даже после отмирания протопласта продолжают выполнять опорную функцию. Различают два типа механических тканей: 1) колленхиму и 2) склеренхиму. Колленхима – механическая ткань молодых растущих органов, возникает очень рано, когда еще продолжается рост органа в длину. Колленхима состоит из живых, вытянутых по оси органа клеток с тупыми или скошенными концами. В клетках часто содержатся хлоропласты. Клеточные стенки утолщены неравномерно и никогда не одревесневают. Граница между первичной и вторичной стенками не выражена, в утолщениях чередуются слои целлюлозы и сильно обводненные слои, богатые пектинами и гемицеллюлозами. В растущем органе стенки клеток должны сохранять способность к растяжению, что возможно только в живых клетках. Функции опорной ткани колленхима может выполнять только в состоянии тургора. Если растение теряет воду, тонкие участки клеточных стенок складываются «гармошкой», побеги теряют упругость и обвисают. Колленхима располагается сразу за покровной тканью в молодых стеблях, цветоносах, черешках листьев, образуя сплошной цилиндр или тяжи в ребрах. В корнях колленхима обычно не встречается. Наиболее характерна колленхима для двудольных, у однодольных растений она встречается редко. В зависимости от характера утолщения стенок клеток различают три типа колленхимы: 1) уголковую, 2) пластинчатую и 3) рыхлую. Уголковая колленхима имеет стенки, утолщенные в углах клеток. Утолщения стенок соседних клеток смыкаются, образуя трех – пятиугольники (рис. 3.21). Уголковая колленхима часто встречается в стеблях травянистых растений, черешках листьев, вдоль главной жилки листа.
Рис. 3.21. Уголковая колленхима на поперечном срезе листового черешка свеклы.
Пластинчатая колленхима имеет утолщения тангенциальных, т. е. параллельных поверхности органа, стенок клеток, которые располагаются параллельными слоями, радиальные стенки остаются тонкими (рис. 3.22). Она встречается, чаще всего, в молодых стеблях древесных растений.
Рис. 3.22. Пластинчатая колленхима на поперечном срезе.
Рыхлая колленхима имеет хорошо выраженные межклетники. Утолщению подвергаются лишь те части стенок, которые прилегают к межклетным пространствам (рис. 3.23). Рыхлая колленхима встречается у некоторых травянистых растений (лопух, дурман). Она сочетает признаки уголковой колленхимы и аэренхимы.
Рис. 3.23. Рыхлая колленхима на поперечном срезе.
Склеренхима встречается наиболее часто, во всех органах: корнях, стеблях, листьях, плодах, цветках, семенах. Клетки склеренхимы имеют равномерно утолщенные и, как правило, одревесневшие стенки. Полость клетки мала, поры простые, щелевидные, немногочисленные. Протопласт, как правило, рано отмирает, и опорную функцию выполняют мертвые клетки. Различают два типа склеренхимы: 1) волокна и 2) склереиды, различающиеся формой клеток. Волокна – прозенхимные клетки, сильно вытянутые в длину и заостренные на концах. Они обеспечивают прочность органов растений на растяжение, сжатие и изгибы. Прочность волокон повышается благодаря тому, что фибриллы целлюлозы проходят в них винтообразно, меняя направление во внешних и внутренних витках. Волокна, расположенные в коровой части осевого органа (во флоэме), называются лубяными. Их длина сильно колеблется: у льна - 40-60 мм, у кендыря – 2-55 мм, а у рами – 350-420 мм. Лубяные волокна с неодревесневающей клеточной стенкой являются ценным сырьем для текстильной промышленности (лен, рами, кенаф). Волокна, находящиеся в древесине (ксилеме), называются древесинными,или волокнами либриформа. Их стенки всегда одревесневшие, длина их не превышает 2 мм. В растениях также часто присутствуют волокна, не связанные с проводящими тканями (листья однодольных). Склереиды – клетки, имеющие различную форму, чаще паренхимную. Они встречаются как поодиночке, в виде идиобластов, так и группами. Это мертвые клетки с очень толстыми одревесневшими стенками, пронизанными поровыми каналами, которые часто ветвятся (рис. 3.24). В зависимости от формы клеток выделяют несколько типов склереид. Наиболее часто встречаются брахисклереиды, или каменистые клетки и астросклереиды. Каменистые клетки имеют более или менее округлую форму (рис. 3.24). Из них состоят косточки вишни, сливы, персика, скорлупа грецкого ореха. Они встречаются в мякоти плодов груши, айвы, рябины, в корнях хрена среди тонкостенных клеток. У груши при созревании плода наблюдается раздревеснение каменистых клеток.
Рис. 3.24. Каменистые клетки околоплодника груши.
Астросклереиды имеют ветвистую форму с отростками, направленными в разные стороны (рис. 3.25). Они располагаются в виде идиобластов в мезофилле листьев некоторых растений (камелия, маслина, кубышка), скрепляя рыхлые ткани подобно шпильке в волосах.
Рис. 3.25. Астросклереида кубышки.
Проводящие ткани Проводящие ткани служат для передвижения по растению растворенных в воде питательных веществ. Они возникли как следствие приспособления растений к жизни на суше. В связи с жизнью в двух средах – почвенной и воздушной, возникли две проводящие ткани, по которым вещества передвигаются в двух направлениях. По ксилеме от корней к листьям поднимаются вещества почвенного питания – вода и растворенные в ней минеральные соли (восходящий, или транспирационный ток). По флоэме от листьев к корням передвигаются вещества, образовавшиеся в процессе фотосинтеза, главным образом сахароза (нисходящий ток). Так как эти вещества представляют собой продукты ассимиляции углекислого газа, транспорт веще ств по флоэме называют током ассимилятов. Проводящие ткани образуют в теле растения непрерывную разветвленную систему, соединяющую все органы – от тончайших корешков до самых молодых побегов. Ксилема и флоэма представляют собой сложные ткани, в их состав входят разнородные элементы – проводящие, механические, запасающие, выделительные. Самыми важными являются проводящие элементы, именно они выполняют функцию проведения веществ. Ксилема и флоэма формируются из одной и той же меристемы и, поэтому, в растении всегда располагаются рядом. Первичные проводящие ткани образуются из первичной латеральной меристемы – прокамбия, вторичные – из вторичной латеральной меристемы – камбия. Вторичные проводящие ткани имеют более сложное строение, чем первичные. Ксилема (древесина) состоит из проводящих элементов – трахеид и сосудов (трахей), механических элементов - древесинных волокон (волокон либриформа) и элементов основной ткани - древесинной паренхимы. Проводящие элементы ксилемы носят название трахеальных элементов. Различают два типа трахеальных элементов – трахеиды и членики сосудов (рис. 3.26). Трахеида представляет собой сильно вытянутую в длину клетку с ненарушенными первичными стенками. Передвижение растворов происходит путем фильтрации через окаймленные поры. Сосуд состоит из многих клеток, называемых члениками сосуда. Членики расположены друг над другом, образуя трубочку. Между соседними члениками одного и того же сосуда имеются сквозные отверстия – перфорации. По сосудам растворы передвигаются значительно легче, чем по трахеидам.
Рис. 3.26. Схема строения и сочетания трахеид (1) и члеников сосуда (2).
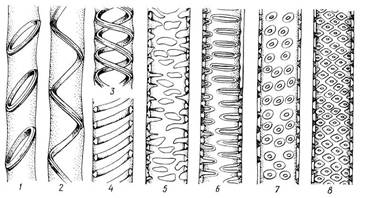
Трахеальные элементы в зрелом, функционирующем состоянии – мертвые клетки, не имеющие протопластов. Сохранение протопластов затрудняло бы передвижение растворов. Сосуды и трахеиды передают растворы не только в вертикальном, но и в горизонтальном направлении в соседние трахеальные элементы и в живые клетки. Боковые стенки трахеид и сосудов сохраняются тонкими на большей или меньшей площади. В то же время они имеют вторичные утолщения, придающие стенкам прочность. В зависимости от характера утолщений боковых стенок трахеальные элементы называются кольчатыми, спиральными, сетчатыми, лестничными и точечно-поровыми (рис. 3.27).
Рис. 3.27. Типы утолщения и поровости боковых стенок у трахеальных элементов: 1 – кольчатое, 2-4 – спиральные, 5 – сетчатое утолщения; 6 – лестничная, 7 – супротивная, 8 – очередная поровость.
Вторичные кольчатые и спиральные утолщения прикрепляются к тонкой первичной стенке посредством узкого выступа. При сближении утолщений и образовании между ними перемычек возникает сетчатое утолщение, переходящее в окаймленные поры. Эту серию (рис. 3.27) можно рассматривать как морфогенетический, эволюционный ряд. Вторичные утолщения клеточных стенок трахеальных элементов одревесневают (пропитываются лигнином), что придает им дополнительную прочность, но ограничивает возможности роста в длину. Поэтому в онтогенезе органа сначала появляются еще способные растягиваться кольчатые и спиральные элементы, не препятствующие росту органа в длину. Когда рост органа прекращается, возникают элементы, неспособные к продольному растяжению. В процессе эволюции первыми появились трахеиды. Они найдены у первых примитивных наземных растений. Сосуды появились значительно позже путем преобразования трахеид. Сосудами обладают почти все покрытосеменные растения. Споровые и голосеменные растения, как правило, лишены сосудов и обладают только трахеидами. Лишь в виде редкого исключения сосуды встречены у таких споровых, как селагинелла, некоторых хвощей и папоротников, а также у немногих голосеменных (гнетовые). Однако у этих растений сосуды возникли независимо от сосудов покрытосеменных. Возникновение сосудов у покрытосеменных растений означало важное эволюционное достижение, так как облегчило проведение воды; покрытосеменные растения оказались более приспособленными к жизни на суше. Древесинная паренхима и древесинные волокна выполняют запасающие и опорные функции соответственно. Флоэма (луб) состоит из проводящих - ситовидных - элементов, сопровождающих клеток (клеток-спутниц), механических элементов – флоэмных (лубяных) волокон и элементов основной ткани – флоэмной (лубяной) паренхимы. В отличие от трахеальных элементов проводящие элементы флоэмы и в зрелом состоянии остаются живыми, а их клеточные стенки – первичными, неодревесневшими. На стенках ситовидных элементов имеются группы мелких сквозных отверстий – ситовидные поля, ч
|
|||||||||
|
|
Последнее изменение этой страницы: 2017-02-05; просмотров: 1640; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.96.159 (0.092 с.) |