Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Регуляторные и контррегуляторные гормоны
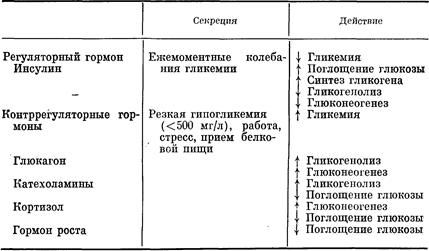
Большое число гормонов, влияющих на обмен энергетических субстратов в организме, можно подразделить на способствующие либо накоплению, либо расходованию энергии. В этом отношении уникальная роль принадлежит инсулину, поскольку он способствует накоплению всех основных видов энергетических веществ (глюкоза, жир и белок). В отличие от него адреналин, глюкагон, кортизол, гормон роста и тиреоидный гормон способствуют расходованию запасов жира, гликогена и/или белка в организме. Эти гормоны можно классифицировать также с точки зрения влияния глюкозы плазмы на их секрецию по принципу обратной связи и с позиций их действия на гликемию. Инсулин — единственный гормон, на секрецию которого влияют нормальные ежемоментные колебания уровня сахара в крови (см. рис. 10—13) и который вызывает снижение последнего. Напротив, каждый из «энергорасходующих» гормонов (за исключением тиреоидного) вызывает повышение уровня сахара в крови. Кроме того, повышение их секреции требует либо явной гипогликемии, либо вмешательства сигналов, не имеющих прямого отношения к изменению уровня глюкозы в крови (например, прием белковой пищи, физическая нагрузка, стресс). В отношении контроля за гликемией инсулин обычно рассматривают как главный регуляторный гормон, тогда как глюкагон, катехоламины, кортизол и гормон роста называют общим термином «контррегуляторные гормоны» [104] (табл. 10—6). Повышающее гликемию действие контррегуляторных гормонов может отражать их прямое влияние на метаболизм гликогена и/или глюконеогенез или влияние, препятствующее инсулиновой стимуляции поглощения глюкозы (см. табл. 10—6). Таблица 10—6. Регуляторные и контррегуляторные гормоны в контроле за гликемией
ВЗАИМОДЕЙСТВИЕ МЕЖДУ ЭНЕРГЕТИЧЕСКИМИ СУБСТРАТАМИ И ГОРМОНАМИ Регуляция обмена веществ в организме в норме основана не только на контролируемых ферментами метаболических путях и разнообразии гормональных сигналов, но и на тонко координированных субстрат-гормональных взаимодействиях. Последние легче всего понять, проанализировав изменения, возникающие в присутствии субстратов (состояние сытости), потребности в субстратах (голодание, физическая работа) или нарушения равновесия субстратов («стрессорная» гипергликемия). Однако перед анализом развивающихся нарушений необходимо рассмотреть нормальный состав тела (т. е. запасы субстратов) и условия при исходном состоянии.
СОСТАВ ТЕЛА Из калорий, входящих в состав типичной диеты американцев или европейцев, 40—45% приходится на долю углеводов, 40% — на долю жира и 15—20% —на долю белка. Однако состав тела человека сильно отличается от состава потребляемой пищи (табл. 10—7). В виде углеводов накапливается незначительное количество калорий. Совместная калорическая ценность гликогена печени (70 г), мышц (200 г) и глюкозы, циркулирующей в крови (2G г), составляет приблизительно 1100 ккал, что существенно ниже общих суточных калорических затрат даже в исходном состоянии. Тем не менее гликоген печени служит важным источником углеводов, идущих на покрытие потребности мозга в исходном состоянии и потребности мышц при работе (см. ниже). С позиций целесообразности ограниченность углеводных запасов оправдана не только их низкой калорической ценностью (4 ккал/г по сравнению с 9 ккал/г для жира), но и необходимостью высокого содержания тканевой воды, входящей в состав запасов гликогена (4 мл/г). Таблица 10—7. Состав тела и энергетические резервы у здорового человека с массой тела 70 кг
Наибольшие резервы энергетических субстратов в организме представлены жиром, накопленным в виде триглицеридов. В отсутствие ожирения запасы жира у человека составляют 20% от общей массы тела и содержат 130000—140000 ккал, т. е. 80% от общего запаса энергетических субстратов в организме. Этого количества калорий могло бы хватить для покрытия исходных потребностей в калориях в течение приблизительно 2 мес. У тучных людей жировая ткань может запасать более 500000 ккал. Независимо от степени ожирения жир несомненно служит наиболее расходуемым и наиболее богатым источником энергетических субстратов для человека.
Основным источником белка в организме является мышечная ткань, в которой накапливается 10 кг (исключая тканевую воду) или 40 000 ккал. В силу роли белка в строении тела, мышечном сокращении и катализе (ферменты), уменьшение его количества на 30—50% оказывается несовместимым с жизнью, несмотря на сохранение остаточной поддающейся мобилизации жировой ткани. Смерть при голодании наступает не от гипогликемии, а от истощения белковых запасов — прекращения функционирования дыхательной мускулатуры, что ведет к терминальной пневмонии. При анализе гомеостатических реакций на потребность в энергетических веществах особую важность приобретает рассмотрение их взаимопревращений в организме. Белки организма (т. е. составляющие их аминокислоты, кроме лейцина) легко превращаются в глюкозу (глюконеогенез). По мере истощения запасов гликогена в печени снабжение углеводами тканей, облигатно зависимых от глюкозы (например, мозг), не прекращается. С другой стороны, жирные кислоты, содержащие четное число углеродных атомов (на долю которых приходится более 95% всех жирных кислот, накопленных в виде триглицеридов), не могут превращаться в глюкозу, так как ткани млекопитающих лишены ферментов, необходимых для глюконеогенеза из ацетил-СоА. Таким образом, в условиях окончательного исчерпания запасов глюкозы (окисление ее в мозге), требующего восполнения ее уровня за счет глюконеогенеза, обязательно происходит истощение белковых запасов. ИСХОДНОЕ СОСТОЯНИЕ Исходное состояние натощак представляет собой переход от состояния сытости к состоянию голодания и может сохраняться в течение 6—12 ч после приема пищи (рис. 10—25). Хотя этот интервал и не отражает стабильного состояния, но тем не менее он является легко установимой точкой отсчета различных отклонений. В исходных условиях из жировой ткани высвобождаются жирные кислоты для удовлетворения энергетических потребностей мышц, сердца и паренхиматозных органов (печень, почки). Дыхательный коэффициент мышцы в покое близок к 0,7, что свидетельствует об окислении ею практически только жира. Углеводы утилизируются главным образом мозгом, в котором окисляется глюкоза до конца со скоростью 125 г/сут. Меньшие количества глюкозы утилизируются покоящейся мышцей и облигатно анаэробными тканями, такими, как форменные элементы крови и мозговой слой почек (рис. 10—26).
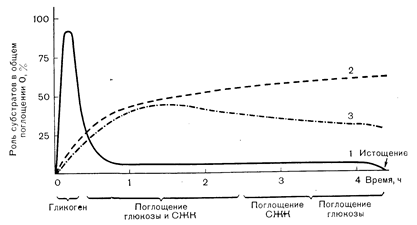
Рис. 10—25. Характер утилизации субстратов мышцами при физической нагрузке различной продолжительности. В первые несколько минут работы главным «топливом», используемым для образования АТФ, служит мышечный гликоген (1). Между 10-й и 40-й минутой работы по 40% всех окисляемых субстратов приходится на долю приносимых с кровью глюкозы и свободных жирных кислот (СЖК). Затем (т. е. при очень длительной работе) происходит прогрессивное увеличение утилизации СЖК (2) и снижение поглощения глюкозы (3) из крови. Хотя мышечный гликоген играет небольшую роль в качестве утилизируемого «топлива» при длительной работе, его полное истощение сопровождается развитием утомления.
Если в утилизации глюкозы принимают участие несколько тканей, то продукция ее ограничена печенью. Поддержание эугликемии зависит от высвобождения глюкозы печенью со скоростью, равной скорости ее утилизации в мозге и периферических тканях вместе (150—250 г/сут, 2—3 мг/кг в 1 мин, 6—10 г/ч). Примерно 15% глюкозы, высвобождаемой после ночного голодания, образуется из гликогена, остальное количество — при глюконеогенезе из лактата, аланина и в меньшей степени пирувата и глицерина [10]. При полном отсутствии продукции глюкозы в печени уровень сахара в крови должен был бы снизиться вдвое в течение 40—60 мин. Поступление жирных кислот в печень приводит к образованию кетоновых тел. Однако скорость кетогенеза в исходном состоянии может обеспечить только не превышающий 0,5 мМ уровень кетокислот в крови.
Гормональным сигналом, позволяющим начаться гликогенолизу, глюконеогенезу и кетогенезу, в исходном состоянии служит снижение уровня инсулина в плазме с 30—100 мкЕД/мл (в состоянии сытости) до 10—20 мкЕД/мл. Таким образом, утилизация
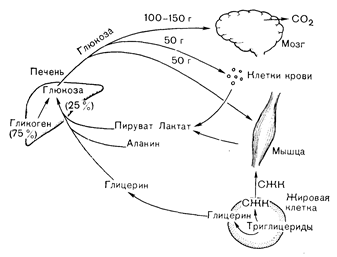
Рис. 10—26. Кругооборот глюкозы в состоянии покоя (после ночного голодания) у здорового человека. Единственным местом продукции глюкозы за счет гликогенолиза (75%) и глюконеогенеза (25%) является печень. Главным местом утилизации глюкозы, потребляющим ее со скоростью 100—150 г/сут, является мозг. Меньшие количества глюкозы поглощаются форменными элементами крови и покоящейся мышцей, служащей источником предшественников глюкозы в виде аланина и интермедиатов гликолиза—лактата и пирувата. Небольшие количества глицерина для глюконеогенеза поставляются адипоцитами. глюкозы в исходном состоянии не зависит от инсулина и почти полностью происходит в мозге. С другой стороны, сохранение исходного Уровня инсулина (и исходной скорости его секреции 0,25—1,5 ЕД/ч) удерживает продукцию глюкозы и кетоновых тел на уровне, не превышающем скорости их утилизации, что предотвращает развитие гипергликемии и гиперкетонемии. При уменьшении скорости секреции инсулина (например, при ювенильном диабете) или в условиях гормональных изменений, препятствующих действию исходного уровня инсулина (см. ниже, раздел «Стрессорная гипергликемия»), концентрация глюкозы и кетоновых тел в крови увеличивается.
|
|||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-26; просмотров: 142; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.224.214.215 (0.006 с.) |