

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Кров рухається повільно, має низхідний напрямок.
СПЕЦКУРС
«ГІСТОФІЗІОЛОГІЯ НЕЙРОЕНДОКРИННОЇ СИСТЕМИ»
Для самопідготовки цього спецкурсу студентам пропонуємо стислий матеріал з ілюстраціями будови залоз.
У середині минулого століття сформувалася нова галузь фізіології - нейроендокринологія, що вивчає структуру і функцію ендокринних елементів нервової тканини, що синтезують нейрогормони, а також вивчає їхню роль в організмі. Однією з центральних проблем нейроендокринології – є дослідження ролі нейросекреторних клітин гіпоталамуса в регуляції функції аденогіпофіза і периферичних ендокринних залоз та вегетативних функцій організму.
У 1963 р. опублікована фундаментальна праця Ернеста і Берти Шаррер «Neuroendocrinology», де викладені основні положення нейроендокринної регуляції функцій ендокринних залоз.
На підставі експериментальних даних зроблено висновки, що в гіпоталамічній ділянці мозку присутні особливі нервові клітини, що здатні до феномену нейросекреції. Нейроцити отримали назву нейросекреторні клітини (НСК), а явище – нейросекреції. Ці нейрони об'єднані в ядра, що лежать поблизу III шлуночку мозку. Нейросекрет має білкову природу – це гіпоталамічні гормони: ліберини (7 видів) та статини (3 види) або нонапептиди (окситоцин і вазопресин).
Між центральними ланками (гіпоталамус, гіпофіз) та ендокринними залозами існують як прямі зв’язки, так і зворотні зв’язки (можуть бути негативні та позитивні).
На сьогодні вважається доведеним:
1. Аденогіпофізотропні нейрогормони (ті, що регулюють функцію аденогіпофіза) виробляються не тільки в гіпоталамусі, але і за його межами; список ідентифікованих аденогіпофізотропних нейрогормонів постійно поповнюється і включає кілька десятків найменувань. Наприклад: тироліберин синтезується як в гіпоталамусі, так і за його межами виконуючи там функцію нейромедіатора.
2. У більшості випадків однакові нейросекреторні нейрони включені до складу не одного, а декількох ядер гіпоталамуса і, навпаки, кожне ядро має гетерогенну популяцію нейронів. Наприклад: СОЯ та ПВЯ містять нейрони, що вироблять окситоцин, вазопресин, тироліберин, кортиколіберин тощо.
3. У нейросекреторних нейронах виробляються не тільки класичні аденогіпофізотропні пептидні і моноамінові нейрогормони, але і гормони, ідентичні за своїми імунологічними характеристиками гормонам аденогіпофіза.
4. Окрім прямого шляху надходження нейрогормонів у портальну систему циркуляції з аксонів до крові, існує й інший, більш складний шлях: з аксонів у III шлуночок мозку і уже відтіля по спеціалізованим епендимним клітинам шлуночка таніцитам у портальну систему циркуляції.
5. Пептидні та моноамінові фізіологічно активні речовини, які синтезуються нейросекреторними нейронами, відіграють роль не тільки нейрогормонів, але і нейротрансмітерів і нейромодуляторів (тобто регулюють передачу нервових імпульсів).
6. В одному нейросекреторному нейроні можуть міститися, а, можливо, і синтезуватися дві і більш фізіологічно активні речовини. Наприклад: нонапептиди та тиреоліберин або нейромедіатор і гормон.
7. Деякі аденогіпофізотропні нейрогормони здатні специфічно впливати на периферичні органи-мішені. Наприклад: люліберин через гормони ЛГ і ФСГ аденогіпофіза впливає на регуляцію функції лише статевих залоз – гонад; кортиколіберин – тільки на синтез гормонів кори наднирників.
8. Вазопресин і окситоцин, окрім їх добре відомого впливу на водно-сольовий обмін і скоротливу здатність гладкої мускулатури ряду органів, також володіють аденогіпофізотропним ефектом, тобто здійснюють потенціюючий вплив на клітини аденогіпофіза.
Зв'язок минулого і сьогодення. Перший завідувач нашої кафедри проф. Перемежко у 1867 р. вперше в гістології звернув увагу на морфологічну гетерогенність різних частин гіпофіза, тобто що він складається з двох частин відомих сьогодні як аденогіпофіз і нейрогіпофіз. Кафедра під керівництвом завідувачів професорів Новикова Б.Г., Гордієнко В.М. і Дзержинського М.Е. вивчає центральні механізми регуляції ендокринних залоз.
Г І П О Т А Л А М У С
Гіпоталамус є вентральним відділом ПРОМІЖНОГО МОЗКУ, межі якого легко встановити за макроорієнтирами на вентральній поверхні головного мозку.
Над гіпоталамусом знаходиться таламус та епіфіз (дорсальна частина проміжного мозку).
На фронтальних зрізах видно, що гіпоталамус розділений щілинним III шлуночком на дві симетричні половини, тобто гіпоталамус утворює стінки III шлуночка мозку та його дно.
Рострально (попереду) гіпоталамус обмежений оптичної хіазмою (перехрест зорових нервів), латерально(з боків)– оптичними трактами і каудально (сзаду) - мамілярними тілами.
СХЕМА БУДОВИ ГІПОТАЛАМУСА ТА ГІПОФІЗА
 | АДЕНОГІПОФІЗ
(передня частка)
| | 
Термін гіпоталамус походить від грецького слова hypothalamus, де hypo - під + thalamos кімната. Синоніми: гіпоталамічна ділянка, сірий бугор.
На свіжому нефіксованому мозку можна виділити 2 ділянки: у центрі – більш темна ділянка, що має вигляд сірого бугра; по периферії – світла, майже прозора ділянка, що включають хіазму і мамілярні тіла.
Сірий бугор – немієліновий гіпоталамус, тобто тут локалізовані тіла НСК (НЕЙРОСЕКРЕТОРНІ КЛІТИНИ). Інша ділянка гіпоталамуса – мієліновий гіпоталамус утворений волокнами (відростки НСК).
Гіпоталамічна ділянка – це парна структура, нейроцити (у кількості приблизно 3000-5000 клітин) якого утворюють гіпоталамічні ядра, що розташовуються симетрично по обидві сторони від III шлуночка. На сьогодні виділяють більше 40 ядер і зон у рострокаудальному, вентродорсальному і латеромедіальному напрямках.
Згідно розповсюдженої класифікації (Сентаготаї й ін.), гіпоталамус поділяється в рострокаудальному напрямку на три відділи: передній (ростральний), середній і задній (каудальний).
Передній гіпоталамус включає такі ядра: СХЯ, СОЯ, переднє гіпоталамічне, ПВЯ і ПЯ.
До складу середнього відділу (медіобазальний гіпоталамус) входять латеральна і дорсальна гіпоталамічні ділянки, АЯ, ВМЯ і ДМЯ.
Задній гіпоталамус включає мамілярні ядра і перифорнікальне ядро. Деякі дослідники до гіпоталамусу відносять також субталамічні утворення, як зона інсерта. |
ГОРМОНИ ГІПОТАЛАМУСА
Ліберини та статини ( див. нижче), що синтезуються в нейроцитах дрібноклітинних ядер гіпоталамуса, попадають у кров і діють на аденогіпофіз завдяки існуванню між гіпоталамусом та гіпофізом портальної воротної системи кровообігу – портальна система гіпофіза. Ця система складається з двох сплетень: первинне капілярне сплетення у зоні серединного підвищення (СП) гіпоталамуса та вторинне капілярне сплетення в аденогіпофізі. Між ними є зв'язок через портальні вени. Кров рухається повільно у низхідному напрямку, тобто від гіпоталамусу до аденогіпофізу.
Нонапептиди( див. нижче) по аксонам СОЯ та ПВЯ, де вони синтезуються, прямують до нейрогіпофізу і виділяють у системний кровоток.
|
| Супраоптичне ядро (СОЯ) розташовано над оптичною хіазмою по латеральному краю гіпоталамуса, доходячи до оптичних трактів. Складається тільки з крупних НСК, що синтезують вазопресин або окситоцин. Нонапептиди збираються у гранули, які містять білок носій та певний нонапептид. Зокрема, окситоцин зв’язаний з нейрофізином I, а вазопресин з нейрофізином II. Секреторні гранули мають діаметр 200 нм, вони оточені мембраною, містять електроннощільний центр. Деякі нейроцити СОЯ синтезують кортиколіберин.
Паравентрикулярне ядро (ПВЯ). Розташовано у стінках III шлуночка мозку над СОЯ, тобто дорсальніше у фронтальній площині. Поділяється на крупноклітинний і дрібноклітинний відділи.
Поліфункціональність ПВЯ пояснюється синтезом у ньому ряду найважливіших нейрогормонів - вазопресина, окситоцина, кортиколіберина, тиреоліберина й інших.
Аксони нейроцитів ПВЯ разом із СОЯ формують так званий супраоптикопаравентрикуло-гіпофізарний тракт,який закінчується у нейрогіпофізі, де відбувається депонування і виділення нонапептидів у загальну системну циркуляцію.
Існує ще один тракт - туберо-інфундибулярний тракт,що утворений аксонами дрібноклітинних ядер гіпоталамуса, які закінчуються на капілярах первинного портального сплетення у СП. Отже так здійснюється вплив ліберинів та статинів гіпоталамуса на залозисті клітини аденогіпофіза.
У цитоплазмі видно нейросекреторну субстанцію, що може включати гормони: окситоцин та вазопресин або інші білкові гормони ліберини або статини, біогенні аміни
ФУНКЦІЯ.НСК гіпоталамуса синтезують гормони, що називають ліберини та статини.Це група гормонів пептидної природи, які далі попадають до портальних судин аденогіпофіза та стимулюють (ліберини) або пригнічують (статини) виділення відповідних тропних гормонів аденогіпофіза. Відомо 7 ліберинів і 3 статини, але структура їх ще не всіх вивчена. Наприклад: тиреоліберин – три пептид (стимулює секрецію ТТГ аденогіпофіза), люліберин – декапептид (стимулює секрецію як ЛГ, так і ФСГ), соматостатин – тетрадекапептид (пригнічує секрецію гормону росту).
У ядрах СОЯ та ПВЯ синтезуються нонапептиди (дев’ять амінокислот): окситоцин і вазопресин, що депонуються у нейрогіпофізі.
Зверніть увагу, що у гіпоталамусі порівняно з іншими ділянками мозку виявлена також найбільша концентрація нейромедіаторів: ДА, НА, А, СЕ та ін.., що свідчить про екстрагіпоталамічні механізми контролю функцій гіпоталамуса.
| Гіпоталамус. Нейроцити СОЯ Забарвлення ПАФ.
 Гіпоталамус. Нейроцити СОЯ.
Гіпоталамус. Нейроцити СОЯ.
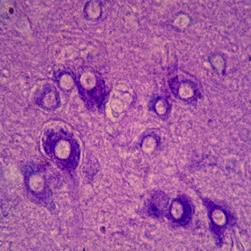 Гіпоталамус. Нейроцити ПВЯ.
Гіпоталамус. Нейроцити ПВЯ.
 Схема будови портальної системи гіпофіза (між гіпоталамусом і гіпофізом)
Схема будови портальної системи гіпофіза (між гіпоталамусом і гіпофізом)
 
1, 2 – ядра СОЯ, ПВЯ, 3 – оптична хіазма, 4 – нервові волокна, 5 – задня частка нейрогіпофіза, 6 – кровоносні судини, 7 - дрібноклітинні ядра, 8 – серединне підвищення, 9 – первинне сплетення капілярів портальної системи, 10 – вторинне сплетення капілярів у складі дистальної частки аденогіпофіза, 11 – портальні вени.
Г І П О Ф І З
| Гіпофіз розташований у гіпофізарній ямці турецького сідла клиноподібної кісти. Турецьке сідло зверху покрите діафрагмою, що є продовженням твердої мозкової оболонки з отвором, через який проходить ніжка гіпофіза, котра пов'язує його з вентральним гіпоталамусом (1) і воронкою мозку (2), що має порожнину (3), яка є продовженням III шлуночку мозку (4). Нижню (звужену) частину воронки називають гіпофізарною ніжкою.
Маса гіпофіза - близько 0,5 г. Гіпофіз поділяють на 2 складові частини.
I. Аденогіпофіз (залозиста частка) складає близько 70% загальної ваги гіпофіза і поділяється на:
а) передня частка (5) – дистальна частка
б) проміжна частка (6) має вид вузької смужки і відокремлена від передньої частки гіпофізарною щілиною (7);
в) туберальная частка (8) - верхній відділ, прилеглий до гіпофізарної ніжки.
II. Нейрогіпофіз включає ділянку СП і гіпофізарну ніжку і задню (нервову) частку гіпофіза (9).
Зовні гіпофіз покритий щільною сполучнотканнинною капсулою (10).
| Гіпофіз кішки
 Гіпофіз людини
Гіпофіз людини

| Передня частка аденогіпофіза
| | | Задня частка нейрогіпофіза
| | 
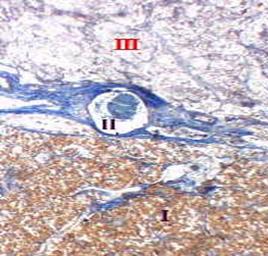
| | На відносно невеликому збільшенні, видно межу між адено- і нейрогіпофізом, тому в полі зору опинилися всі три частки гіпофіза:
передня частка (I), дуже вузька проміжна частка (II) аденігофіза і задня частка (III) нейрогіпофіза.
Ядра всіх клітин забарвлюються в оранжевий колір, а колагенові волокна - в синій.
Ось тому, видно, що в передній частці переважають клітини, в проміжній частці - прошарки сполучної тканини, а в задній частці - відносно мало і клітин, і строми.
| Гіпофіз людини. Забарвлення сумішшю Маллорі по Генденгайну.
| | | | | | | Передня частка гіпофіза (дистальна частка аденогіпофіза)
| Серед тканинних елементів передньої частки розрізняють:
секреторні клітини (1) - клітини залозистого епітелію, сполучнотканинна строма (2) численні синусоїдальні (тобто дуже широкі) капіляри (3).
Секреторні клітини лежать групами формуючи тяжі (трабекули), що утворюють розгалужену мережу.
Строма утворена прошарками пухкої сполучної тканини, які розділяють трабекула
Синусоїдальні капіляри складають вторинне капілярне сплетення портальної системи гіпофіза, що забезпечує вплив ліберинів і статинів гіпоталамуса на клітини-"мішені" аденогіпофіза.
Гормони гіпоталамуса спочатку попадають у кров через первинне капілярне сплетення,на стінках якого закінчуються аксони НСК гіпоталамуса.
Обидва сплетення утворюють портальну систему гіпофіза.
| Аденогіпофіз людини. Забарвлення сумішшю Маллорі за Генденгайном
 Аденогіпофіз птахів
Аденогіпофіз птахів


| Залозисті клітини передньої частки аденогіпофіза
Залозисті клітини розрізняються по тінкторіальним властивостях (здатності забарвлюватись певним барвником).
Тінкторіальні властивості обумовлені присутністю в цитоплазмі ендокриноцитів секреторних гранул з білковим гормоном.
Якщо в цьому гормоні переважають кислі функціональні групи, цитоплазма клітини є базофільною, а коли переважають основні групи, цитоплазма – ацидофільна.
Якщо гранул практично немає зовсім, клітина виявляється хромофобною (тобто є незабарвленою).
АДЕНОГІПОФІЗ ЛЮДИНИ
У складі однієї трабекули присутні клітини різних типів. Проте, є ділянки, де переважають ті або інші клітини.
Хромофобні(4), тобто клітини, що не сприймають барвник і тому при даному забарвленні світло-сірі; складають приблизно 60 % всіх клітин. З них 2-5% припадає на клітини з часточковими ядрами (на знімку не помітні);
Хромофільні – базофільні клітини двох типів (5.А – 5.Б), що забарвлюються у фіолетовий колір; 5-10 % всіх клітин; і ацидофільні клітини (6) з оранжевою цитоплазмою, 30-35 %

| з переважанням ацидофілів (оранжева цитоплазма)

| з переважанням базофілів (фіолетова цитоплазма)

| | На даному препараті видно переважно ацидофільні клітини (6).
Хоча в невеликій кількості присутні інші клітини -хромофобні (4) і базофільні (5).
Поряд є прошарки сполучної тканини (1) та синусоїдальні капіляри (2) і в яких еритроцити (3).
| Тут, навпаки, - декілька трабекул з відносно високим вмістом базофільних клітин (5.А-5.Б). Але видно інші залозисті клітини - зокрема, хромофобні (4).
Зверніть увагу на те, що в одних базофільних клітинах (5.А) ядро знаходиться приблизно в центрі, а в інших (5.Б) - різко ексцентрично, тоді як в центрі розташовується світла пляма.
|
ХРОМОФОБНІ КЛІТИНИ
| Морфологія
| Функція
| | Клітини невеликі за об'ємом. В більшості випадків (за винятком кортикотропоцитов) цитоплазма не містить секреторних гранул, тому погано сприймає барвники. Отже дані клітини дрібні та світло-сірі.
| а) стовбурові клітини;
б) базофільні та ацидофільні клітини, які ще не накопичили специфічні гранули або позбулися їх в результаті інтенсивної секреції;
в) а також кортикотропні клітини (синтезують АКТГ).
| | Примітка: кортикотропоцити нерідко відносять до особливого типу хромофільних клітин, що не є ні базофільними, ні ацидофільними.
| | Характеристика кортикотропоцитов.
Кортикотропоцити відрізняються неправильною формою клітини і лапатим ядром (1). Крім того, вони містять гормональні гранули (2): у вигляді мембранних пухирців з щільною білковою серцевиною.
| 
| | | | |
ХРОМОФІЛЬНІ КЛІТИНИ
Е П І Ф І З
| (Локалізація епіфізу (шишковидної залози):
I. поперечний зріз мозку;
II. вигляд зверху, мозолисте тіло і свод видалені; судинна покришка третього шлуночку розітнута і відтягнута в сторони.
| | Епіфіз (1) розташований на даху (2) середнього мозку, під мозолистим тілом (3).
Його вага - всього лише близько 0,25 г, тобто приблизно удвічі менше, ніж за гіпофіз.
| 
|
Будова епіфіза
| Епіфіз містить 4 основних компоненти:
Сполучнотканинна строма (1.А - Б),
пінеалоцити (2.А - Б) - секреторні клітини, гліальні клітини (3), чисельні капіляри (4).
Від капсули (1.А) відходять і перегородки (1.Б), що розділяють залозу на часточки.
| Епіфіз людини: мале (I) і велике (II) збільшення. /

|
| З віком епіфіз піддається інволюції. Йде нагромадження мозкового піску (вуглекислий, фосфорнокислий Ca2+, Mg2+) - 1. Водночас збільшується кількість строми.
Виділяють декілька типів будови епіфіза.: целюлярний, трабекулярний і альвеолярний.
Відкладення солей кальцію - мозковий пісок.
| |  
Клітини епіфіза
| Пінеалоцити
| Пінеалоцити мають досить великий розмір, ядра бульбашковидні, від клітин відходять чисельні відростки, що контактують з капілярами.
| | Разновідості пінеалоцитів
| Розрізняють дві різновидності пінеалоцитів - світлі клітини (2.А) - з невеликим вмістом секреторних гранул, темні клітини (2.Б) - з високим вмістом гранул. Мабуть, одні і ті ж клітини можуть бути як в одному, так і в іншому стані
| | Гормони
| У темряві - мелатонін з вираженим антигонадотропним ефектом, тоді як в інший час доби - ряд інших регуляторів функції гіпофіза і периферичних залоз (серотонін, тиреоліберин тощо), різні нейромедіатори.
| | Глія
| Гліальні клітини (3) розташовуються між пінеалоцитами і мають дрібні розміри, щільне ядро,і як і пінеалоцити мають відростки.
|
Щ И Т О В И Д НА З А Л О З А
| Щитовидна залоза найбільша з ендокринних залоз. Її вага близько 30-40 г.
Розташована попереду трахеї.
Включає дві бічні частки (1) перешийок (2) і у 30-50 % випадків також має вузьку пірамідальну частку (3).
Покрита зовні сполучнотканинною капсулою. Від останньої углиб залози відходять прошарки, що ділять її на часточки.
| Рисунок щитовидної залози

| Гістологічна будова
Структурною одиницею щитовидної залози є фолікули та інтерфолікулярні острівці.
Фолікули (1) вистелені тиреоїдним епітелієм і заповнені гомогенним колоїдом (2).
Клітини носять назву тироцити або тиреоцити
Невелика частка залозистих клітин формує компактні скупчення поза фолікулами – це так званий  екстрафолікулярний епітелій (3).
На препараті лівіше знаходиться паращитовидна залоза (I) екстрафолікулярний епітелій (3).
На препараті лівіше знаходиться паращитовидна залоза (I)
| Щитовидна залоза. Забарвлення гематоксилін-еозином.

| Різні види тироцитів:
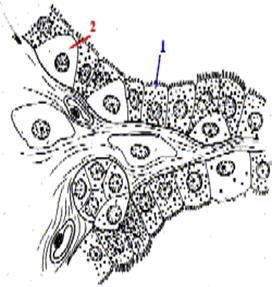
Як у стінках фолікулів, так і поза ними присутні клітини двох видів (при звичайній мікроскопії майже невиразні). Диференціювати їх можна за поглинанням радіоактивного йоду (чорні гранули на рис.):
одні клітини - фолікулярні (1)- поглинають йод (заповнені чорними гранулами) – тиреоцити А
 інші - парафолікулярні клітини (2) - не поглинають йод (тому на рис. вони світлі) – тиреоцити С.
існують ще тироцити В –так звані клітини Ашкіназі-Гюртле, онкоцити тощо. інші - парафолікулярні клітини (2) - не поглинають йод (тому на рис. вони світлі) – тиреоцити С.
існують ще тироцити В –так звані клітини Ашкіназі-Гюртле, онкоцити тощо.
| Диференціація клітин щитовидної залози за поглинанням йоду.
| | | | |
Розвиток щитовидної залози
| ПАРАФОЛІКУЛЯРНІ
ТИРОЦИТИ С
| | | ФОЛІКУЛЯРНІ КЛІТИНИ – ТИРОЦИТИ А,
ЩО УТВОРЮЮТЬ СТІНКИ ФОЛІКУЛІВ
| | | ЕПІТЕЛІАЛЬНИЙ ВИРОСТ, ЯКИЙ РОСТЕ
ВЗДОВЖ ГЛОТКИ
ВЗДОВЖ ГЛОТКИ
| | | ВИП’ЯЧУВАННЯ СТІНКИ ГЛОТКИ
| | 
Отже, фолікулярні клітини мають епітеліальне походження, а парафолікулярні клітини - нервове.
|
Види секреторних клітин
| Розрізняють два види секреторних клітин (3): головні паратироцити (з базофільною цитоплазмою) і
оксифільні паратироцити.
біля кровоносних судин зустрічаються паратироцити, що синтезують кальцитонін
Окрім паратироцитів (3) видно також прошарки пухкої сполучної тканини (1) і капіляри (4).
| Паращитовидна залоза. Забарвлення гематоксилін-еозином.
Велике збільшення

|
Головні паратироцити
| Гормон
| Паратгормон (гіперкальцимічний фактор) підвищує вміст Са2+ в крові.
| | Тетанія
| При видаленні паращитовидних залоз розвивається тетанія - судоми, обумовлені різким зниженням концентрації в крові іонів Са2+. Механізм тетанії пов'язаний з тим, що Са2+ за умов норми зменшує збудливість м'язових і нервових мембран, але при недостатності Са2+ в крові збудливість м’язів зростає.
| | Разновідності паратироцитів
| Поміж головних паратироцитів розрізняють світлі і темні клітини. Мабуть це пов'язано з різним вмістом в клітинах секреторних гранул, тобто чим їх більше, тим клітина виглядає темніше.
|
Оксифільні паратироцити
| За розмірами крупніші, ніж головні клітини; мають оксифільну цитоплазму. З’являються в онтогенезі пізніше головних клітин. Припускають, що ці клітини є одним з видів функціонального стану головних паратироцитів. За іншою версією, в них утворюється антагоніст паратгормона - кальцитонін, що знижує вміст Са2+ в крові.
|
Н А Д Н И Р Н И К И
| Наднирники (1) - парні органи, розташовані над верхніми кінцями нирок (2) в зачеревній клітковині.
Вага двох наднирників близько 8 г.
| 
|
Гістологічна будова
| Зовні кожний наднирник покритий сполучнотканинною капсулою (1). Під нею в органі розрізняють два шари.
| Наднирники кішки. Забарвлення гематоксилін-еозином.

| | Кіркова речовина (2.А-2.В)
| Знаходиться з поверхні; на свіжому розрізі має жовтуватий колір через велику кількість ліпідів.
| | Мозкова речовина (3)
| Зосереджена в середині органу; на розрізі - темніше і м'якше.
| | Ці шари наднирників розглядують як дві відносно самостійні залози, які продукують різні по структурі і функції гормони, але які об'єднані в єдиний орган із спільним кровопостачанням.
У нижчих хребетних тварин відповідні залози існують роздільно.
|
Кора наднирників
| У корі секреторні клітини називаються кірковими ендокриноцитами або частіше спонгіоцитами, тому що мають піняву цитоплазму ніби то губчасту (Губки – Spongia).
У корі можна виділити 3 зони (2.А, 2.Б, 2.В), які відрізняються розташуванням епітеліальних клітин та синтезом відповідного стероїдного гормону і механізмом регуляції.
| 
| Зведена таблиця даних по корі наднирників
| Зона кори
| Зовнішній вигляд
| Гормони
| Регуляція
| | Клубочкова (2.А)
| Знаходиться на поверхні; спонгіоцити утворюють округлі скупчення (клубочки).
| Мінералкортикоїди:
альдостерон
|  Ренін (нирки)
ангіотензиноген Ренін (нирки)
ангіотензиноген
 (печінка)
ангіотензини (кров) (печінка)
ангіотензини (кров)
| | Пучкова (2.Б)
| Найширша по товщині; має вид довгих пучків, орієнтованих перпендикулярно поверхні капсули.
| Глюкокортикоїди:
кортикостерон
кортизол
гідрокортизон
| АКТГ
(аденогіпофіз)
| | Сітчаста
(2.В)
| Найглибша зона кори; за розмірами ближче до клубочкової. Клітини утворюють пухку сітку
| Андрогенита естрогени
| АКТГ викликає проліферацію клітин всіх зон кори
|
Клубочкова зона кори
| Наднирники мають товсту сполучнотканинну капсулу (1), від якої відходять тонкі прошарки (2) углиб залози. У капсулі і прошарках знаходяться кровоносні судини.
Поверхнева ділянка кори наднирників, що безпосередньо лежить під капсулою – це клубочкова зона кори.
Ширина цієї зони коливається від 50 до 70 мкм, а об’єм складає 10-15 % об’єму всієї кори.
Спонгіоцити утворюють округлі скупчення - клубочки (4.А), у складі яких нараховується до 5-6 великих кубічних чи полігональних клітин із дрібнозернистою ацидофільною цитоплазмою і великим ядром.
При забарвленні на ліпіди (суданом III) в цитоплазмі клітин виявляється менше ліпідних включень, ніж у двох інших зонах корки, але у зв'язку з синтезом стероїдів ці клітини (як і в інших зонах кори) мають розвинену гладку ЕПС.
Над і під клубочковой зоною знаходяться дві вузькі росткові ділянки з дрібних малодиференційованих клітин. За рахунок клітин однієї з них (5) поповнюються пучкова і сітчаста зони.
| Надниркова. Капсула і клубочкова зона кори. Забарвлення гематоксилін-еозином.


|
Пучкова зона кори
| Перехід клубочкової зони в пучкову відбувається поступово. При цьому спостерігається переорієнтація епітеліальних тяжей і зміни внутрішньої структури кліток.
Пучкова зона ширше інших зон і складає в середньому 63,5-67,5% від об’єму всієї кори, а середня ширина цієї зони варіює від 400 до 500 мкм. Ця зона представлена тяжами спонгіоцитів, розташованих перпендикулярно до капсули і до клубочкової зони. Ці клітни мають кубічну, призматичну, іноді полігональну і навіть кулясту форму.
Пучки розділені сполучно тканин-
ними прошарками (6) в яких розташовуються кровоносні капіляри (3).
| Наднирники. Пучкова зона. Забарвлення гематоксилін-еозином.Середнє збільшення.

| | Спонгіоцити мають піняву (губчасту) цитоплазму - із-за наявності великої кількості крупних ліпідних включень глюкокортикоїдів. Ось тому їх називають світлими. До того ж за розміром вони більше за клітини сусідніх зон кори; мають добре розвинену гладку ЕПС (де відбувається синтез стероїдів), а кристи мітохондрій - тубулярного типу.
Зустрічаються також темні клітини: у них секреторні гранули ще не встигли накопичитися.
| Велике збільшення

|
Сітчаста зона кори
| Сітчаста зона коризнаходиться на кордоні кори з мозковою речовиною. Має вигляд анастомозуючих тяжів (пухка сітка). Її ширина складає 100-150 мкм, тобто вона ширше клубочкової, але значно уступає пучковій зоні.
У цій зоні клітини дрібніше, ніж у пучкової, і відрізняються поліморфністю. Клітини тут округлої, бокаловидної і полігональної форми і контактують відразу з декількома капілярами. Мітохондрії численні, але їх менше, ніж у клітинах пучкової зони, вони мають змішану будову крист (тубулярно-везикулярні).
| Наднирники. Сітчаста зона.
Забарвлення гематоксилін-еозином.

|
Мозкова речовина
Мозкова речовина знаходиться в центрі наднирників, відношення її маси до маси кори - 1:3. На кордоні кори і мозкової речовини знаходиться тонка внутрішня капсула 2-3 мкм.
Залозисті клітини мозкової речовини (8) - мозкові ендокриноцити, або хромафінні клітини. Клітини називаються хромафінними тому, що вони здатні при обробці біхроматом відновлювати його до нижчих оксидів хрому, який відкладається в клітинах у вигляді бурого осаду. Подібні клітини виявляються і в інших частинах організму і є видозміненими нейронами симпатичної нервової системи. Між хромафінними клітинами є багато синусоїдальних капілярів і крупні венули. Вони синтезують адреналін і норадреналін.
Хромафінні клітини зібрані в скупчення від 2 до 6 клітин (діаметр 25-30 мкм), мають циліндричну, кубічну і полігональну форми з дрібнозернистою цитоплазмою і великим світлим ядром.
Розрізняють епінефроцити (адреноцити) і норепінефроцити (норадреноцити). Вважають, що світліші клітини синтезують адреналін, а темніші - норадреналін. Гормони концентруються у чисельних мембранних пухирцях (секреторних гранулах) з щільною серцевиною, до складу якої входить спеціальний білок-носій, що акумулює гормони.
Адреноцити містять більш великі гранули 0,6 мкм, норадреноцити – більш дрібні 0,1 мкм. Ці гранули мають характерну будову.

С Т А Т Е В І З А Л О З И
| С І М ’ Я Н И К
Паренхіма сім’яника поділена на I50-250 часточок. У цих часточках лежать по одному звивистому сім’яно-му канальцю, а також кровоносні судини та інтерстиціальна тканина, котра представлена клітинами Лейдіга (синтезують гормон тестостерон, що переважно виділяється у лімфу).
Стінка звивистого сім’яного канальця складається із сполучнотканинної власної оболонки, в якій розрізняють: внутрішній шар - базальна мембрана має глікопротеїдну природу, над нею розташований волокнистий шар з колагенових і еластичних волокон, потім міоїдний шар, утворений декількома рядами міоїдних клітин. Скорочення міоїдних клітин обумовлює перистальтику звитих насінних канальців і просування. по них ще нерухомих сперматозоїдів.
На базальній мембрані лежать сперматогонії та клітини Сертолі (1 ряд), далі верхні шарі складаються з статевих клітин на різних стадіях сперматогенезу: сперматоцити I і II порядку (2-4 ряди), сперматиди (5-6 ряди) та сперматозоїди, що своїми хвостами обернені у просвіт канальцю.
Клітини Сертолі виконують опорно-трофічну функцію. Також беруть участь в утворенні гематотестикулярного бар’єру; здатні до фагоцитозу статевих клітин, що загинули та залишків, які відокремились від сперматид; секретують рідину, в яку попадають сперматозоїди в просвіті канальців; мають рецептори до ФСГ та тестостерону у зв’язку з цим виконують координуючу функцію при сперматогенезі; синтезують гормон інгібін (пригнічує продукцію ФСГ аденогіпофізом), фактор стимулюючий поділ сперматогоній і андрогензвя’зуючий білок (тільки при зв’язування з ним тестостерон діє на сперматиди).
| Звивистий сім’яний канадець, зб.х40
 Звивистий сім’яний канадець,
зб.х90
Звивистий сім’яний канадець,
зб.х90


|
Я Є Ч Н И К
|



