
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Схема 3. Биосинтетич. и др. р-ции, сопутствующие циклу трикарбоновых к-т; аминокислоты, отмеченные звездочкой, в организме высших животных несинтезирую гея.Содержание книги
Поиск на нашем сайте
Сукцинил-КоА в р-ции 5, катализируемой сукцинил-КоА -синтетазой, подвергается распаду, в результате к-рого энергия тиоэфирной связи сукцинил-КоА запасается в виде синтезир. нуклеозидтрифосфата (у бактерий, грибов, растений-АТФ, у животных - ГТФ). В р-ции 6, катализируемой сукцинатдегидрогеназой, происходит превращение сукцината в фумарат. Фермент входит в состав более сложного сукцинатдегидрогеназного комплекса (комплекса II) дыхат. цепи, поставляя восстановит. эквиваленты, образующиеся в р-ции, в дыхат. цепь. Фумарат-гидратаза, катализирующая р-цию 7, осуществляет гидратирование фумарата с образованием L-малата. Активность фермента ингибируется АТФ. Последняя р-ция цикла, 8, катализируется L-малатдегид-рогеназой; L-малат при этом превращ. в оксалоацетат, к-рый может взаимод. с новой молекулой ацетил-КоА. Р-ция обратима, равновесие сдвинуто в сторону образования L-малата (DG0 +29,73 кДж/моль), но в условиях функционирования цикла конечным продуктом р-ции является оксалоацетат. Предполагают, что катализируемые индивидуальными ферментами р-ции осуществляются благодаря действию надмолекулярного "сверхкомплекса", т. наз. метаболона. Преимущества такой организации ферментов очевидны -при этом не происходит диффузии кофакторов и субстратов, что способствует более эффективной работе цикла. Наличие в трикарбоновых кислот цикле 4 восстановит. р-ций (3, 4, 6 и 8), в результате к-рых на 1 молекулу CH3C(O)SKoA синтезируются 3 молекулы НАДН и 1 молекула ФАДН, определяет необходимые условия для его функционирования. Непрерывная работа цикла требует реокисления НАДН и ФАДН, к-рое в аэробных условиях, как правило, осуществляется через совокупность переносчиков электронов, составляющих дыхат. цепь, и сопровождается запасанием значит. кол-ва энергии. У животных это 11 молекул АТФ на 1 молекулу окисленного ацетил-КоА. У грибов, растений и особенно бактерий число молекул АТФ, образующихся при окислении НАДН и ФАДН, м.б. меньше вследствие разветвления дыхат. цепи. В анаэробных условиях вместо трикарбоновых кислот цикла функционируют его окислит. ветвь до 2-оксоглутарата (р-ции 1: 2: 3) и восстановительная - от оксалоацетата до сукцината (р-ции 8: 7: 6). При этом не происходит запасания большого кол-ва энергии и ф-ция цикла целиком определяется доставкой в-в для синтеза клеточного материала. При переходе организма от покоя к активному состоянию возникает потребность в ускоренной мобилизации энергии (обменных процессов). У животных это достигается, в частности, шунтированием наиб. медленных р-ций трикарбоновых кислот цикла (р-ции 1-3) и преимуществ. окислением сукцината. При этом исходный субстрат укороченного трикарбоновых кислот цикла (2-оксоглутарат) образуется в результате быстрой р-ции переаминирования: Глутамат + Оксалоацетат Связь трикарбоновых кислот цикла с глиоксилатным циклом осуществляется благодаря синтезу в последнем сукцината, к-рый в трикарбоновых кислот цикле окисляется до оксалоацетата и служит, т. обр., поставщиком молекул с 4 атомами С в оба цикла на начальных стадиях их функционирования. Благодаря этому возможно функционирование этих циклов, когда в организме избыток соед. с 2 атомами С, напр. при выращивании бактерий на средах с СН3СООН и С2Н5ОН, а также при прорастании семян масличных растений, во время к-рого усиленно образуется ацетил-КоА. Др. модификация трикарбоновых кислот цикла (т. наз. 4-аминобутиратный шунт) - превращение 2-оксоглутарата в сукцинат, через глу-таминовую к-ту, 4-аминобутират и янтарный семиальдегид (3-формилпропионовая к-та). Эта модификация имеет большое значение для ткани мозга, в к-рой ок. 10% глюкозы распадается по этому пути. Тесное сопряжение р-ций трикарбоновых кислот цикла с дыхат. цепью, особенно в митохондриях животных, а также особенности регуляции активности индивидуальных ферментов цикла (для большинства из них АТФ является ингибитором) предопределяют снижение активности цикла в условиях генерирования высокого фосфорильного потенциала (отношения АТФ/АДФ) в клетке, и наоборот - активацию цикла при пониж. фосфорильном потенциале. У большинства растений, бактерий и мн. видов грибов тесное сопряжение цикла с дыхат. цепью преодолевается развитием альтернативных несопряженных путей окисления, позволяющих поддерживать дыхат. активность и активность трикарбоновых кислот цикла на высоком уровне даже в условиях высокого фосфорильного потенциала. Трикарбоновых кислот цикл открыт в 1937 X. Кребсом и У. Джонсоном. ПЕНТОЗОФОСФАТНЫЙ ЦИКЛ (пентозный путь, гексо-зомонофосфатный шунт, фосфоглюконатный путь), совокупность обратимых ферментативных р-ций, в результате к-рых происходит окисление глюкозы до CO2 с образованием восстановленного никотинамидадениндинуклеотид-фосфата (НАДФН) и H +, а также синтез фосфорилир. Сахаров, содержащих от 3 до 7 атомов С. Пентозофосфатный цикл осуществляется в цитозоле (жидкой фазе) клеток животных, растений (особенно в темноте) и микроорганизмов. У растений часть р-ций пентозофосфатного цикла участвует также в образовании гексоз при фотосинтезе.
|
||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 164; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.11 (0.007 с.) |

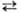 2-Оксоглутарат + Аспартат
2-Оксоглутарат + Аспартат


