Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Фармакология синаптической передачиСодержание книги Похожие статьи вашей тематики
Поиск на нашем сайте
ФАРМАКОЛОГИЯ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ В ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЕ То, что мы думаем, гораздо менее сложно, чем то, чем мы думаем. Станислав Лем Периферической нервной системой (ПНС) называют совокупность образований, лежащих вне головного и спинного мозга: корешки мозга, нервные стволы и их сплетения, нервные узлы и рецепторы. Периферическая нервная система состоит из 2 отделов: · Афферентного (чувствительного) – по волокнам этого отдела импульсы от рецепторов кожи, слизистых оболочек и исполнительных органов поступают в ЦНС. · Эфферентного (двигательного) – по волокнам этого отдела импульсы от ЦНС поступают ко всем внутренним органам.
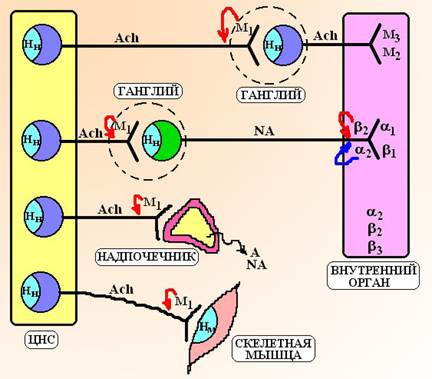
В эфферентном отделе ПНС выделяют соматическую и вегетативную (автономную) часть. Соматическая часть нервной системы находится под контролем сознания и управляет такими функциями, как движение, дыхание, поддержание позы тела. Медиатором в соматических проводниках является ацетилхолин. Тела соматических нейронов располагаются в ядрах черепных нервов и передних рогах спинного мозга, а их отростки нигде не прерываясь поступают к скелетным мышцам. Таким образом, эфферентные соматические проводники состоят из 1 нейрона. Вегетативная часть нервной системы не подчиняется контролю нашего сознания, т.е. она является непроизвольной. Вегетативная нервная система контролирует жизненно важные функции внутренних органов и обеспечивает жизнедеятельность организма. Вегетативные проводники состоят из 2 нейронов и подходят ко всем внутренним органам. В зависимости от анатомического строения вегетативную систему подразделяют на 2 части: 1. Парасимпатическую часть. Тела первых нейронов этого отдела располагаются в ядрах черепных нервов: III пары (n. oculomotorius), VII (n. facialis), IX (n. glossofaringeus) и X пары (n. vagus) – это, так называемый, краниальный отдел и в боковых рогах серого вещества сакрального отдела спинного мозга (S2-S4). Отростки этих нейронов (преганглионарные волокна) покидают спинной мозг и направляются к нервным узлам (ганглиям), которые расположены вблизи иннервируемых органов или в толще их стенок. Здесь импульс передается на тело второго нейрона и по его отросткам (постганглионарным волокнам) он поступает к исполнительным органам. Таким образом, преганглионарные волокна парасимпатического отдела длиннее, чем постганглионарные (обычно соотношение их длины составляет 1-2:1, а у блуждающего нерва 8.000:1). Медиатором как в преганглионарных, так и в постганглионарных нейронах является ацетилхолин. 2. Симпатическую часть. Тела первых нейронов симпатического отдела вегетативной системы располагаются в боковых рогах серого вещества тораколюмбального отдела спинного мозга (C8-L3). Отростки этих нейронов (преганглионарные волокна) направляются к ганглиям, которые расположены паравертебрально (по бокам от позвоночного столба) или превертебрально (кпереди от позвоночного столба). В ганглиях импульс переходит на тело второго нейрона. Отростки второго нейрона (постганглионарные волокна) покидают ганглий и направляются к эффекторным органам. Таким образом, в симпатической системе преганглионарные волокна короче, чем постганглионарные (обычно соотношение их длины 1:20). Медиатором в преганглионарных волокнах является ацетилхолин, а в постганлионарных волокнах – норадреналин. В ряде органов строение симпатического отдела вегетативной системы отличается от классического варианта: · Мозговое вещество надпочечников иннервируется преганглионарными симпатическими нервами, которые содержат в качестве медиатора ацетилхолин. Ганглиев, где бы происходило переключение на второй нейрон, здесь не имеется. Мозговое вещество надпочечников развивается из нервной трубки эмбриона, поэтому его можно рассматривать как аналог ганглия в котором произошла атрофия постганглионарных нервных волокон, а медиатор норадреналин трансформировался в гормон адреналин[1]. · Сосуды почек иннервируются постганглионарными симпатическими нервами в которых роль медиатора выполняет дофамин. · Терморегуляционные потовые железы кожи и сосуды скелетных мышц получают симпатическую иннервацию, в которой постганглионарные волокна содержат в качестве медиатора ацетилхолин.
Схема 2. Вегетативная и соматическая иннервация. Ach – ацетилхолин, NA – норадреналин. Как правило, внутренние органы получают двойную иннервацию – симпатическую и парасимпатическую, однако, имеются некоторые исключения из этого общего правила. Гладкие мышцы бронхов получают главным образом парасимпатическую иннервацию, функции симпатической иннервации берет на себя гормон адреналин. Гладкие мышцы сосудов, напротив, получают исключительно симпатическую иннервацию не имея парасимпатических проводников (хотя рецепторы, характерные для парасимпатического отдела вегетативной системы, могут присутствовать на эндотелиальных клетках). В стенке органов ЖКТ располагаются крупные нейрональные сети: межмышечная (сплетение Ауэрбаха) и подслизистая (сплетение Мейснера). Эти сети иногда рассматривают как особую часть вегетативной системы – метасимпатический отдел. Метасимпатическая часть вегетативной системы получает эфферентные сигналы от преганглионарных парасимпатических волокон и постганглионарных симпатических нейронов, а также афферентные импульсы от чувствительных нейронов стенки кишки. На уровне метасимпатической системы происходит интеграция поступающих импульсов и формируется интегральный сигнал, который координирует работу органа (например, синхронизирует сокращение стенок кишки и одновременное раскрытие ее сфинктеров для продвижения содержимого). Таблица 1. Сравнительная характеристика отделов вегетативной нервной системы.
Таблица 2. Сравнительная характеристика холинорецепторов клетки.
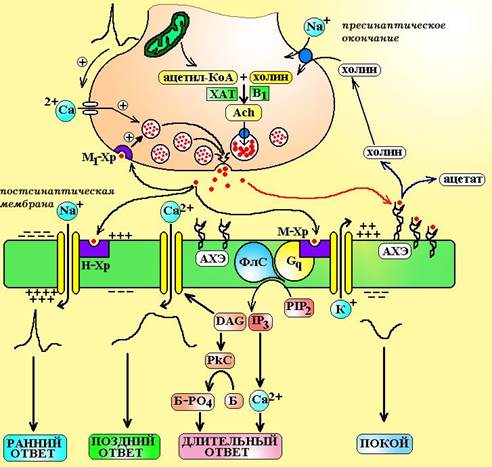
Примечание: a-бунгаротоксин – яд тайваньской гадюки Bungaris multicintus и кобры Naja naja. PTMA – фенилтриметиламмоний DMPP – диметилфенилпиперазин HHSDP – гексагидросиладифенол АВУ – атриовентрикулярный узел САУ – синоаурикулярный узел Синтез ацетилхолина проводит особый фермент холинацетилтрансфераза, путем ацетилирования холина. Образовавшийся ацетилхолин поступает в везикулы при помощи антипортера переносчика в обмен на протон. Работа этого переносчика может быть заблокирована векзамиколом. Обычно в каждой везикуле содержится от 1.000 до 50.000 молеукл ацетилхолина, а общее число везикул в пресинаптическом окончании достигает 300.000. 2. Выделение медиатора. Во время фазы покоя, через пресинаптическую мембрану выделяются единичные кванты медиатора (изливается содержимое 1 везикулы). Одна молекула ацетилхолина способна вызвать изменение потенциала мембраны всего на 0,0003 мВ, а то количество, которое содержится в 1 везикуле – на 0,3-3,0 мВ. Такие миниатюрные сдвиги не вызывают развития биологического ответа, но поддерживают физиологическую реактивность, тонус ткани-мишени. Активация синапса происходит в тот момент, когда на пресинаптическую мембрану приходит потенциал действия. Под влиянием потенциала мембрана деполяризуется и это вызывает открытие воротного механизма медленных кальциевых каналов. По этим каналам ионы Са2+ поступают в пресинаптическое окончание и взаимодействуют с особым белком в мембране везикул – синаптобревином (VAMP). Синаптобревин переходит в активированное состояние и начинает выполнять роль своеобразного «крючка» или якоря. Этим якорем везикулы фиксируются к пресинаптической мембране в тех местах, где лежат особые белки – SNAP-25 и синтаксин-1. В последующем эти белки инициируют слияние мембраны везикул с мембраной аксона и выталкивают медиатор в синаптическую щель подобно поршню насоса. При прохождении потенциала действия через пресинаптическую мембрану одновременно опустошаются 2.000-3.000 везикул.
Схема 4. Передача сигнала в холинергическом синапсе. ХАТ – холинацетилтрансфераза, В1 – тиамин, Ach – ацетилхолин, М1-Хр – М1-холинорецепторы, АХЭ – ацетилхолинэстераза, ФлС – фосфолипаза С, PIP2 – фосфатидилинозитол бифосфат, IP3 – инозитол трифосфат, DAG – диацилглицерол, PkC – протеинкиназа С, Б – белок-фермент, Б-РО4 – фосфорилированная форма белка-фермента. Процесс выделения медиатора может быть нарушен под влиянием ботулотоксина (токсин бактерий Clostridium botulinum). Ботулотоксин вызывает протеолиз белков, участвующих в выделении медиатора (SNAP-25, синтаксин, синаптобревин). a-латротоксин – яд паука «черная вдова» связывается с белком SNAP-25 (нейрексином) и вызывает спонтанный массивный экзоцитоз ацетилхолина. 3. Развитие биологического ответа. В синаптической щели путем диффузии ацетилхолин поступает к постсинаптической мембране, где активирует холинорецепторы. При взаимодействии с Н-холинорецепторами происходит открытие натриевых каналов и на постсинаптической мембране генерируется потенциал действия. В том случае, если ацетилхолин активирует М-холинорецепторы, сигнал передается через систему G-белков на фосфолипазу С, ионные каналы К+ и Са2+ и все это приводит в конечном итоге к изменению поляризации мембраны, процессов фосфорилирования внутриклеточных белков. Помимо постсинаптической мембраны ацетилхолин может воздействовать на холинорецепторы пресинаптической мембраны (М1 и М2). При активации ацетилхолином М1-пресинаптического рецептора выделение медиатора усиливается (положительная обратная связь). Роль М2-холинорецепторов на пресинапетической мембране недостаточно ясна, полагают, что они могут тормозить секрецию медиатора. Развитие биологического ответа можно вызвать при введении лекарственных веществ, которые стимулируют холинорецепторы или предотвратить, если ввести средства, блокирующие эти рецептры. Повлиять на развитие эффекта можно и не затрагивая рецепторы, а воздействуя лишь на пострецепторные механизмы: · Токсин коклюшной палочки может активировать Gi-белок и снижать активность аденилатциклазы на затрагивая М-холинорецептор; · Токсин холерного вибриона может активировать Gs-белок и повышать активность аденилатциклазы; · Дитерпен форсколин из растения Coleus forskohlii способен непосредственно активировать аденилатциклазу в обход рецепторов и G-белков. 4. Окончание действия медиатора. Время существования ацетилхолина в синаптической щели составляет всего 1 мС, после чего он подвергается гидролизу до холина и остатка уксусной кислоты. Уксусная кислота быстро утилизируется в цикле Кребса. Холин в 1.000-10.000 раз менее активен, чем ацетилхолин, 50% его молекул подвергаются обратному захвату в аксон для ресинтеза ацетилхолина, остальная часть молекул включается в состав фосфолипидов. Гидролиз ацетилхолина осуществляет особый фермент – холинэстераза. В настоящее время известно 2 его изоформы: · Ацетилхолинэстераза (AChE) или истинная холинэстераза – осуществляет высокоспецифичный гидролиз ацетилхолина и локализуется на постсинаптической мембране холинергических синапсов. · Бутирилхолинэстераза (ButChE) или псевдохолинэстераза – осуществляет низкоспецифичный гидролиз эфиров. Локализуется в плазме крови и перисинаптическом пространстве. Сравнительная характеристика этих ферментов представлена в таблице 3. Таблица 3. Сравнительная характеристика холинэстераз.
Таблица 4. Сравнительная харктеристика адренорецепторов.
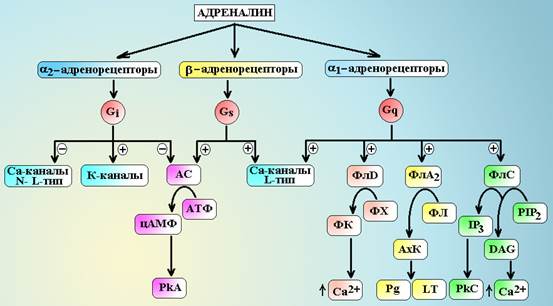
Примечание: A – адреналин, NA – норадреналин, Iso – изопреналин, МПС – мочеполовая система. a1-адренорецепторы посредством Gq-белка передают сигнал на несколько эффекторных систем: ] Фосфолипазу С, которая гидролизует фосфатидилинозитол бифосфат (PIP2) до инозитов трифосфата (IP3) и диацилглицерола (DAG). Молекулы IP3 вызывают выход ионов Са2+ из внутриклеточного депо и активируют зависимые от Са2+ ферменты (кальмодулин). DAG – обеспечивает активацию протеинкиназы С и фосфорилирование внутриклеточных белков, а также открывает Са2+-каналы мембраны. Под влиянием ионов Са2+ и активного кальмодулина происходит дефосфорилирование киназы легких цепей миозина и она переходи в активную нефосфорилированную форму, при этом начинают фосфорилироваться легкие цепи миозина и запускается процесс сокращения гладкомышечных клеток (см. схему 6.). ] Фосфолипазу А2, которая гидролизует фосфолипиды с выделение арахидоновой кислоты. В последующем арахидоновая кислота трансформируется в простагландины и лейкотриены. ] Фосфолипазу D, которая гидролизует фосфатидилхолин до фосфатидной кислоты. Молекулы фосфатидной кислоты вызывают выделение ионов Са2+ из депо, активируют АДФ-рибозилирующий фактор. ] Показана возможность активации G-белками Са2+-каналов клетки. a2-адренорецепторы посредством Gi-белка также передают сигнал на несколько эффекторных систем: ] Gi-белок снижает активность аденилатциклазы и уменьшает синтез цАМФ в клетке. В итоге, активность зависимых от цАМФ протеинкиназ падает. ] Через G0-белки тормозятся Са2+-каналы L- и N-типов. ] bg-субъединицы G-белка активируют К+-каналы мембраны. ] Относительно недавно было обнаружено, что bg-субъединицы Gi-белка могут стимулировать митоген-активирующие протеинкиназы (МАРК), которые обеспечивают процессы деления и размножения стволовых клеток.
Схема 5. Передача сигнала с адренорецепторов. АС – аденилатциклаза, PkA – протеинкиназа А, PkC – протеинкиназа С, ФлС – фосфолипаза С, ФлА2 – фосфолипаза А2, ФлD – фосфолипаза D, ФХ – фосфатидилхолин, ФЛ – фосфолипиды, ФК – фосфатидная кислота, АхК – арахидоновая кислота, PIP2 – фосфатидилинозитол бифосфат, IP3 – инозитол трифосфат, DAG – диацилглицерол, Pg – простагландины, LT – лейкотриены. b-адренорецепторы всех типов реализуют свое действие через Gs-белки. a-субъединицы этого белка активируют аденилатциклазу, которая обеспечивает синтез в клетке цАМФ из АТФ и активацию цАМФ зависимой протеинкиназы А. bg-субъединицы Gs-белка активируют Са2+-каналы L-типа и т.н. maxi-K+-каналы. Под влиянием цАМФ-зависимой протеинкиназы А происходит фосфорилирование киназы легких цепей миозина и она переходит в неактивную форму, не способную фосфорилировать легкие цепи миозина. Процесс фосфорилирования легких цепей прекращается и гладкомышечная клетка расслабляется. Таблица 5. Фармакологическая модуляция синаптической передачи, не связанная с воздействием на циторецепторы.
Таблица 6. Эффекты стимуляции симпатических и парасимпатических проводников.
Таблица 7. Характеристика отдельных медиаторов неадренергического ФАРМАКОЛОГИЯ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-12-30; просмотров: 543; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.27 (0.012 с.) |

 Схема 1. Афферентный и эфферентный отделы нервной системы.
Схема 1. Афферентный и эфферентный отделы нервной системы.