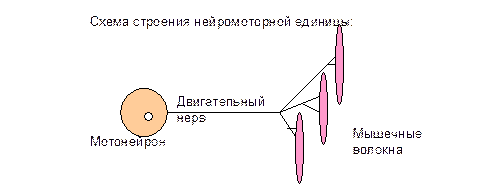Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Восходящие проводниковые путиСтр 1 из 3Следующая ⇒
МЕТОДИЧЕСКИЕ УКАЗАНИЯ ДЛЯ СТУДЕНТОВ ПРИ ПОДГОТОВКЕ К ПРАКТИЧЕСКОМУ ЗАНЯТИЮ
Киев – 2013 1.Актуальность темы: Спинной мозг в ЦНС является первым уровнем регуляции физиологичных функций, и в частности, двигательных функций. Именно здесь расположены нервные центры, на которые осуществляют свои влияния нервные центры головного мозга, передавая информацию нисходящими ведущими путями и обеспечивая приспосбительные реакции организма. В то же время нервные центры головного мозга получают информацию восходящими путями о состоянии регулируемых параметров, которое обеспечивает коррекцию параметров приспосбительной реакции. В случае поражения проводящих путей спинного мозга, которые обеспечивают его связи с центрами спинного мозга, характер спинальных рефлекторных реакций изменяется, или они совсем не осуществляются при повреждении спинальных центров. Знание механизмов функционирования нейронов спинного мозга, его проводниковой и рефлекторной функции играет большую роль в профессиональной деятельности врача-стоматолога, потому что помогает ему понять основные механизмы регуляции работы челюстно-лицевого аппарата человека в норме и патологии, что необходимо в его повседневной работе. 2.Учебные цели: Ø Делать выводы о состоянии двигательных функций организма - мышечного тонуса, позы, локомоций, двигательных рефлексов после поперечного перереза мозга на разных уровнях ЦНС и повреждения двигательных структур Ø Анализировать регулируемые параметры и механизмы активации рецепторов как следящих устройств, при осуществлении двигательных рефлексов. Ø Делать выводы о состоянии двигательных рефлексов, которые замыкаются на разных уровнях ЦНС, и строении их рефлекторных дуг. Ø Делать выводы о роли проводящих путей ЦНС в обеспечении сенсорных и двигательных функций организма.
3.Базовый уровень подготовки Студенты имеют навыки из предыдущих учебных дисциплин:
4.Задание для самостоятельной работы во время подготовки к практическому занятию 4.1.Перечень основных терминов, параметров, характеристик, которые должен усвоить студент при подготовке к практическому занятию
4.2. Теоретические вопросы 1) Понятие о позе тела и условиях ее обеспечения. Установочные позные и двигательные рефлексы организма. 2) Виды сенсорной информации, которые поступают в спинной мозг от рецепторов организма. 3) Рефлекторная деятельность спинного мозга, виды спинальных рефлексов. Тонические и фазические двигательные рефлексы спинного мозга, их физиологичное значение и строение рефлекторных дуг. Эффективность тонических двигательных рефлексов спинного мозга при автономной его деятельности. 4) Зависимость спинальных рефлексов от деятельности центров головного мозга. Спинальный шок.
5) Медиальная и латеральная нисходящие системы регуляции двигательных функций организма. Характер влияний каждого из путей этих систем на спинальные а-мотонейроны разного функционального значения. Способы связи волокон нисходящих путей со спинальными а-мотонейронами. Практические работы 1) Исследование спинального шока у лягушки. 2) Исследование мышечного тонуса спинальной лягушки. 3) Исследование позы спинальной лягушки. 4) Исследование фазических кожно-мышечных двигательных рефлексов спинальной лягушки. 5) Исследование сухожильных рефлексов у человека. Содержание темы Регуляция двигательных функций при участии центральной нервной системы (ЦНС) - это регуляция: тонуса скелетных мышц; позы тела; перемещение частей тела относительно туловища; перемещение тела в пространстве - локомоций. Разные отделы ЦНС осуществляют регуляцию двигательных функций, влияя на нейромоторные единицы.
Количество мышечных волокон в нейромоторных единицах разное - от нескольких единиц до десятков, сотен, тысяч, что связано с регуляцией силы сокращения мышц и их функцией. Мышечные волокна нейромоторных единиц принадлежат к двум группам: а) быстрые, б) медленные. Быстрые мышечные волокна - имеют длительность одиночного сокращения около 20-50 мс - могут сокращаться в анаэробных условиях - развивают значительную силу - быстро утомляются. Медленные мышечные волокна - имеют длительность одиночного сокращения 100-200 мс - сокращаются в аэробных условиях - развивают меньшую силу - долго не утомляются. Скелетные мышцы человека по строению смешанные - в их состав входят как быстрые, так и медленные мышечные волокна. Мотонейроны расположены в передних рогах спинного мозга, среди них есть два вида: 1.Малые мотонейроны: иннервируют малое количество мышечные волокон; имеют низкие пороги и активируются самыми первыми; генерируют ПД с небольшой частотой, которая приводит к меньшей силе сокращения мышечных волокон. 2. Большие мотонейроны: иннервируют значительное количество мышечных волокон; имеют высокие пороги и активируются позже; генерируют ПД с большей частотой, которая приводит к большей силе сокращения мышечных волокон.
А. Сенсорная функция спинного мозга: центральный анализ информации от рецепторов. Спинной мозг получает информацию от проприорецепторов о состоянии опорно-двигательной системы и от экстерорецепторов кожи - о контакте с внешней средой, на основании анализа этой информации он обеспечивает осуществление спинальных двигательных рефлексов. К проприорецепторам принадлежат: а. Мышечные веретена - расположены параллельно мышечным волокнам и передают информацию по афферентным нервным волокнам группы Аa об изменениях длины мышц (статические изменения) и скорости их растяжения (динамические изменения). б. Сухожильные рецепторы Гольджи - расположены в сухожилиях, имеют высокие пороги и передают информацию по афферентным нервным волокнам группы Аb о растяжении сухожилий при сокращении мышц. в. Тельца Пачини - расположены в мышцах, передают информацию по афферентным нервным волокнам группы В о вибрации, подобные им проприорецепторы расположены в суставных сумках и связках и передают информацию об изменениях движения в суставах (скорость движения и величину угла).
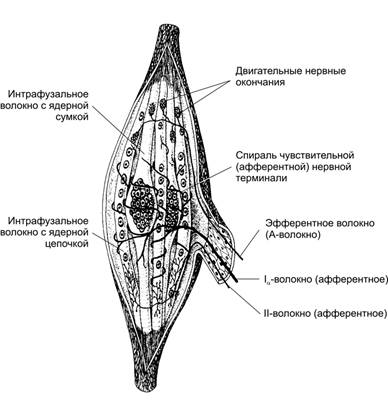
г. Свободные нервные окончания - расположены в мышцах, они высоко пороговые, принадлежат к ноцицепторам - рецепторам повреждения, передают информацию по афферентным нервным волокнам группы В, С. Мышечные веретена, их строение. Их количество в разных скелетных мышцах разное. Основными структурными элементами мышечного волокна являются интрафузальные мышечные волокна, нервные волокна и капсула.
Схема строения мышечного веретена. Интрафузальные мышечные волокна (МВ) с компактным скоплением ядер — волокна с ядерной сумкой, в интрафузальных МВ с ядерной цепочкой ядра распределены по длине волокна более равномерно. К веретену подходят афферентные и эфферентные нервные волокна. Аннулоспиральные (первичные) сенсорные окончания образованы безмиелиновыми терминалями афферентных Ia‑волокон в экваториальной зоне обоих типов интрафузальных МВ. Ближе к концам интрафузальных МВ (чаще МВ с ядерной цепочкой) расположены терминали тонких афферентных II-волокон — вторичные окончания. Эфферентные Аg-волокна образуют нервно-мышечные синапсы с интрафузальными МВ в концевой их части. цепочкой. Ефферентни Аg-волокна образуют нервно-м’язові синапсы из інтрафузальними МВ в их дистальной части. Мышечные волокна. Мышечное веретено содержит от 1 до 10 коротких интрафузальных мышечных волокон. В средней (экваториальной) их части ядра образуют компактное скопление (волокна с ядерной сумкой) или располагаются цепочкой (волокна с ядерной цепочкой). Нервные волокна. Терминали Ia‑волокон образуют спираль в пределах экваториальной зоны обоих типов интрафузальных МВ (первичные, или аннулоспиральные окончания). Терминали более тонких II-волокон заканчиваются на интрафузальных МВ в области, расположенной рядом с экваториальной (вторичные окончания, чаще встречаются в МВ с ядерной цепочкой). Эфферентные Аg‑волокна образуют нервно-мышечные синапсы с интрафузальными МВ в концевой их части Капсула. Комплекс интрафузальных МВ с нервными терминалями окружён многослойной капсулой, наружные слои которой являются производными периневрия, а внутренние рассматриваются как аналоги эндоневрия. При растяжении мышц растягиваются интрафузальные мышечные волокна и передают информацию от спиралевидных рецепторов по афферентным нервным волокнам к а-мотонейронам спинного мозга, активация а-мотонейронов приводит к сокращению екстрафузальных мышечных волокон - уменьшению их длины.
Гамма - мотонейроны получают информацию от разных структур головного мозга и передают нервные импульсы к интрафузальным мышечным волокнам, которые сокращаются и растягивают их ядерную сумку, обеспечивая тем самым возможность передачи информации во время сокращения мышц к а-мотонейронам и другим отделам ЦНС. Альфа-мотонейроны и гамма-мотонейроны активируются одновременно сигналами от двигательных структур головного мозга (коактивация), что позволяет поддерживать растянутыми мышечные веретена во время сокращения мышц и передавать все время информацию о динамических и статических изменениях мышц. При растяжении мышц растягиваются интрафузальные мышечные волокна и передают информацию от спиралевидных рецепторов по афферентным нервным волокнам к а-мотонейронам спинного мозга, активация а-мотонейронов приводит к сокращению экстрафузальных мышечных волокон - уменьшению их длины.
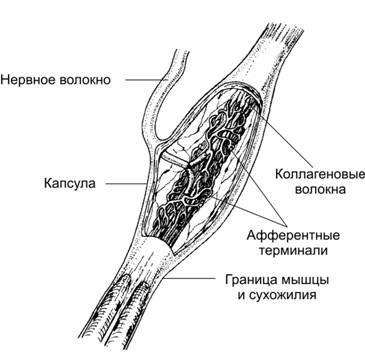
Сухожильные рецепторы Гольджи расположены в сухожилиях на границе с мышцой. Рецептор имеет веретенообразное строение и окружен капсулой. Терминали афферентных нервных миелиновых волокон расположены среди пучков коллагеновых волокон, которые расположены в заполненном жидкостью пространстве. Рецепторы активируются при растяжении сухожилия мышцей, которая сокращается, что приводит к рефлекторному расслаблению этой мышцы.
Схема структуры сухожильного рецептора Гольджи.
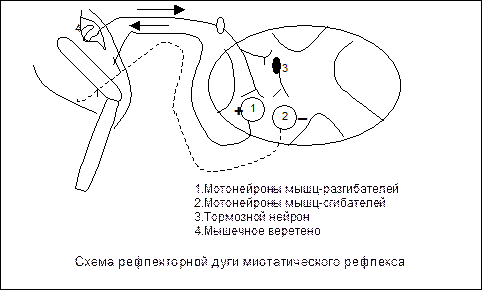
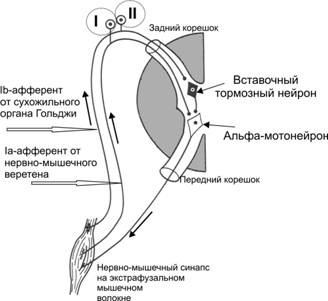
В. Рефлекторна функция спинного мозга. Спинальные двигательные рефлексы. В центрах спинного мозга замыкаются рефлекторные дуги двигательных рефлексов: а) тонических, что лежат в основе регуляции тонуса мышц; б) фазических, что обеспечивают перемещение частей тела относительно туловища. Спинальные рефлексы являются сегментарными или межсегментарными. Рефлексы растяжения с мышечных веретен – миотатические:
- возникают при растяжении мышечных веретен в результате растяжения мышц; - информация от мышечных веретен (первичных спиралеподобных нервных окончаний) по афферентным нервным волокнам передается к а-мотонейронам спинного мозга, которые активируются; - активация а-мотонейронов приводит к передаче информации к скелетным мышцам по эфферентним нервным волокнам, следствием этого является их сокращение; - альфа-мотонейроны мышц-антагонистов тормозятся; - рефлекторная дуга миотатичеких рефлексов, которые возникают при растяжении мышечных веретен, является моносинаптической; - при отсутствии нисходящих активирующих влияний двигательных центров головного мозга на а-мотонейроны и гамма-мотонейроны миотатические рефлексы поддерживают мышечный тонус, но он является ослабленным. 2) Рефлексы растяжения с рецепторов сухожилий: - возникают при растяжении сухожилий мышцей, которая сокращается;
- информация от рецепторов сухожилий Гольджи передается по афферентным нервным волокнам к тормозным нейронам спинного мозга, которые вызывают постсинаптическое торможение а-мотонейронов мышцы, которая сокращалась, следствием этого является ее расслабление. Рефлекторная дуга рефлекса является полисинаптической.
Схемы рефлекторных дуг: 1) рефлекса растяжения (І) с мышечных веретен (что приводит к сокращению той же мышцы; 2) тормозного рефлекса растяжения с рецепторов сухожилий Гольджи (II), которая имеет тормозной нейрон, который приводит к торможению а-мотонейрона и расслаблению мышцы, которая сокращалась. 3) Флексорные кожно-мышечные рефлексы: - возникают при раздражении рецепторов кожи -информация передается по афферентным волокнам к альфа-мотонейронам спинного мозга, которые активируются; - от альфа-мотонейронов нервные импульсы передаются по эфферентным нервным волокнам к мышцам-флексорам, которые сокращаются, следствием чего есть сгибание конечности и отдаление ее от раздражителя; - рефлекторные дуги кожно-мышечных рефлексов являются полисинаптическими.
С. Проводниковая функция спинного мозга. Проводящие пути образованы цепями нейронов, которые соединены между собой синапсами, и обеспечивают передачу информации от нейрону к нейрону, от ядра к ядру. Материалы для самоконтроля 6.1. Дайте ответы на вопрос 1) Какое значение имеет тот факт, что в процессе эволюции животных все большая часть волокон нисходящих путей не непосредственно заканчивается в спинном мозге на а-мотонейронах ( прямая моносинаптическая связь), а связывается с ними через интернейроны (прямая полисинаптическая связь)? 2) Какие из нисходящих путей активируют преимущественно флексорные, а какие – экстензорные мотонейроны спинного мозга? Чем можно объяснить тот факт, что в процессе эволюции большинство нисходящих путей стали преимущественно активировать именно флексорные мотонейроны? 3) Одинаковая ли длительность спинального шока у лягушки и мартышки? О чем это свидетельствует? 4) Как и почему изменится тонус мышц животных при удалении головного мозга? Как это влияет на способность животных сохранить антигравитационную позу? Выберите правильный ответ 1.В эксперименте на лягушке сделали поперечный разрез между продолговатым и спинным мозгом. Какой характер мышечного тонуса и какая поза будет наблюдаться у лягушки?
2.У животного в эксперименте перерезали задние корешки спинного мозга, следствием чего стали изменения в зоне иннервации задними корешками:
3.Какой из приведенных путей активирует спинальные мотонейроны мышц-сгибателей:
4. У животного в эксперименте перерезали передние корешки спинного мозга, следствием чего стали изменения в зоне иннервации задними корешками:
5. Какой из приведенных путей активирует спинальные мотонейроны мышц-разгибателей:
6.У человека после спинномозговой травмы отсутствует болевая и температурная чувствительность. Какие проводниковые пути спинного мозга повреждены?
7.У спинальной лягушки перерез седалищного нерва привел к увеличению длины задней конечности на стороне перереза, что стало следствием повреждения рефлекторной дуги одного из рефлексов
8.У человека обнаружили изменения характера коленного рефлекса, которые стали следствием повреждения локального спинального нервного центра в одном из отделов спинного мозга:
9.В ответ на сильное быстрое сокращение мышцы возникает ее рефлекторное расслабление, которое является следствием раздражения одного из рецепторов:
10.В эксперименте раздражали электрическим током гамма-мотонейроны, которые иннервируют дистальные концы интрафузальных мышечных волокон мышечных веретен, которые расположены в икроножной мышце, следствием чего стало:
Описание практических работ Литература Основная 1. Физиология человека (учебник для медвузов). Покровский В.М., Коротько Г.Ф., ред. М.: Медицина, 2003, 2005. 2. Физиология. Под ред. В.Г.Шевчука. Пособие по физиологии.- Винница: Новая книга. 2005. – 576 с. (русский перевод практических работ кафедрой). 3. Методические указания для самостоятельной работы студентов. Утверждены на методическом заседании кафедры. 2006 Дополнительная
2.Физиология человека: в 3-х томах. Перевод с английского. Под ред. Р.Шмидта и Г.Тевса. – М: Мир, 1996
МЕТОДИЧЕСКИЕ УКАЗАНИЯ ДЛЯ СТУДЕНТОВ ПРИ ПОДГОТОВКЕ К ПРАКТИЧЕСКОМУ ЗАНЯТИЮ
Киев – 2013 1.Актуальность темы: Спинной мозг в ЦНС является первым уровнем регуляции физиологичных функций, и в частности, двигательных функций. Именно здесь расположены нервные центры, на которые осуществляют свои влияния нервные центры головного мозга, передавая информацию нисходящими ведущими путями и обеспечивая приспосбительные реакции организма. В то же время нервные центры головного мозга получают информацию восходящими путями о состоянии регулируемых параметров, которое обеспечивает коррекцию параметров приспосбительной реакции. В случае поражения проводящих путей спинного мозга, которые обеспечивают его связи с центрами спинного мозга, характер спинальных рефлекторных реакций изменяется, или они совсем не осуществляются при повреждении спинальных центров. Знание механизмов функционирования нейронов спинного мозга, его проводниковой и рефлекторной функции играет большую роль в профессиональной деятельности врача-стоматолога, потому что помогает ему понять основные механизмы регуляции работы челюстно-лицевого аппарата человека в норме и патологии, что необходимо в его повседневной работе. 2.Учебные цели: Ø Делать выводы о состоянии двигательных функций организма - мышечного тонуса, позы, локомоций, двигательных рефлексов после поперечного перереза мозга на разных уровнях ЦНС и повреждения двигательных структур Ø Анализировать регулируемые параметры и механизмы активации рецепторов как следящих устройств, при осуществлении двигательных рефлексов. Ø Делать выводы о состоянии двигательных рефлексов, которые замыкаются на разных уровнях ЦНС, и строении их рефлекторных дуг. Ø Делать выводы о роли проводящих путей ЦНС в обеспечении сенсорных и двигательных функций организма. 3.Базовый уровень подготовки Студенты имеют навыки из предыдущих учебных дисциплин:
4.Задание для самостоятельной работы во время подготовки к практическому занятию 4.1.Перечень основных терминов, параметров, характеристик, которые должен усвоить студент при подготовке к практическому занятию
4.2. Теоретические вопросы 1) Понятие о позе тела и условиях ее обеспечения. Установочные позные и двигательные рефлексы организма. 2) Виды сенсорной информации, которые поступают в спинной мозг от рецепторов организма. 3) Рефлекторная деятельность спинного мозга, виды спинальных рефлексов. Тонические и фазические двигательные рефлексы спинного мозга, их физиологичное значение и строение рефлекторных дуг. Эффективность тонических двигательных рефлексов спинного мозга при автономной его деятельности. 4) Зависимость спинальных рефлексов от деятельности центров головного мозга. Спинальный шок. 5) Медиальная и латеральная нисходящие системы регуляции двигательных функций организма. Характер влияний каждого из путей этих систем на спинальные а-мотонейроны разного функционального значения. Способы связи волокон нисходящих путей со спинальными а-мотонейронами. Практические работы 1) Исследование спинального шока у лягушки. 2) Исследование мышечного тонуса спинальной лягушки. 3) Исследование позы спинальной лягушки. 4) Исследование фазических кожно-мышечных двигательных рефлексов спинальной лягушки. 5) Исследование сухожильных рефлексов у человека. Содержание темы Регуляция двигательных функций при участии центральной нервной системы (ЦНС) - это регуляция: тонуса скелетных мышц; позы тела; перемещение частей тела относительно туловища; перемещение тела в пространстве - локомоций. Разные отделы ЦНС осуществляют регуляцию двигательных функций, влияя на нейромоторные единицы.
Количество мышечных волокон в нейромоторных единицах разное - от нескольких единиц до десятков, сотен, тысяч, что связано с регуляцией силы сокращения мышц и их функцией. Мышечные волокна нейромоторных единиц принадлежат к двум группам: а) быстрые, б) медленные. Быстрые мышечные волокна - имеют длительность одиночного сокращения около 20-50 мс - могут сокращаться в анаэробных условиях - развивают значительную силу - быстро утомляются. Медленные мышечные волокна - имеют длительность одиночного сокращения 100-200 мс - сокращаются в аэробных условиях - развивают меньшую силу - долго не утомляются. Скелетные мышцы человека по строению смешанные - в их состав входят как быстрые, так и медленные мышечные волокна. Мотонейроны расположены в передних рогах спинного мозга, среди них есть два вида: 1.Малые мотонейроны: иннервируют малое количество мышечные волокон; имеют низкие пороги и активируются самыми первыми; генерируют ПД с небольшой частотой, которая приводит к меньшей силе сокращения мышечных волокон. 2. Большие мотонейроны: иннервируют значительное количество мышечных волокон; имеют высокие пороги и активируются позже; генерируют ПД с большей частотой, которая приводит к большей силе сокращения мышечных волокон.
А. Сенсорная функция спинного мозга: центральный анализ информации от рецепторов. Спинной мозг получает информацию от проприорецепторов о состоянии опорно-двигательной системы и от экстерорецепторов кожи - о контакте с внешней средой, на основании анализа этой информации он обеспечивает осуществление спинальных двигательных рефлексов. К проприорецепторам принадлежат: а. Мышечные веретена - расположены параллельно мышечным волокнам и передают информацию по афферентным нервным волокнам группы Аa об изменениях длины мышц (статические изменения) и скорости их растяжения (динамические изменения). б. Сухожильные рецепторы Гольджи - расположены в сухожилиях, имеют высокие пороги и передают информацию по афферентным нервным волокнам группы Аb о растяжении сухожилий при сокращении мышц. в. Тельца Пачини - расположены в мышцах, передают информацию по афферентным нервным волокнам группы В о вибрации, подобные им проприорецепторы расположены в суставных сумках и связках и передают информацию об изменениях движения в суставах (скорость движения и величину угла). г. Свободные нервные окончания - расположены в мышцах, они высоко пороговые, принадлежат к ноцицепторам - рецепторам повреждения, передают информацию по афферентным нервным волокнам группы В, С. Мышечные веретена, их строение. Их количество в разных скелетных мышцах разное. Основными структурными элементами мышечного волокна являются интрафузальные мышечные волокна, нервные волокна и капсула.
Схема строения мышечного веретена. Интрафузальные мышечные волокна (МВ) с компактным скоплением ядер — волокна с ядерной сумкой, в интрафузальных МВ с ядерной цепочкой ядра распределены по длине волокна более равномерно. К веретену подходят афферентные и эфферентные нервные волокна. Аннулоспиральные (первичные) сенсорные окончания образованы безмиелиновыми терминалями афферентных Ia‑волокон в экваториальной зоне обоих типов интрафузальных МВ. Ближе к концам интрафузальных МВ (чаще МВ с ядерной цепочкой) расположены терминали тонких афферентных II-волокон — вторичные окончания. Эфферентные Аg-волокна образуют нервно-мышечные синапсы с интрафузальными МВ в концевой их части. цепочкой. Ефферентни Аg-волокна образуют нервно-м’язові синапсы из інтрафузальними МВ в их дистальной части. Мышечные волокна. Мышечное веретено содержит от 1 до 10 коротких интрафузальных мышечных волокон. В средней (экваториальной) их части ядра образуют компактное скопление (волокна с ядерной сумкой) или располагаются цепочкой (волокна с ядерной цепочкой). Нервные волокна. Терминали Ia‑волокон образуют спираль в пределах экваториальной зоны обоих типов интрафузальных МВ (первичные, или аннулоспиральные окончания). Терминали более тонких II-волокон заканчиваются на интрафузальных МВ в области, расположенной рядом с экваториальной (вторичные окончания, чаще встречаются в МВ с ядерной цепочкой). Эфферентные Аg‑волокна образуют нервно-мышечные синапсы с интрафузальными МВ в концевой их части Капсула. Комплекс интрафузальных МВ с нервными терминалями окружён многослойной капсулой, наружные слои которой являются производными периневрия, а внутренние рассматриваются как аналоги эндоневрия. При растяжении мышц растягиваются интрафузальные мышечные волокна и передают информацию от спиралевидных рецепторов по афферентным нервным волокнам к а-мотонейронам спинного мозга, активация а-мотонейронов приводит к сокращению екстрафузальных мышечных волокон - уменьшению их длины. Гамма - мотонейроны получают информацию от разных структур головного мозга и передают нервные импульсы к интрафузальным мышечным волокнам, которые сокращаются и растягивают их ядерную сумку, обеспечивая тем самым возможность передачи информации во время сокращения мышц к а-мотонейронам и другим отделам ЦНС. Альфа-мотонейроны и гамма-мотонейроны активируются одновременно сигналами от двигательных структур головного мозга (коактивация), что позволяет поддерживать растянутыми мышечные веретена во время сокращения мышц и передавать все время информацию о динамических и статических изменениях мышц. При растяжении мышц растягиваются интрафузальные мышечные волокна и передают информацию от спиралевидных рецепторов по афферентным нервным волокнам к а-мотонейронам спинного мозга, активация а-мотонейронов приводит к сокращению экстрафузальных мышечных волокон - уменьшению их длины.
Сухожильные рецепторы Гольджи расположены в сухожилиях на границе с мышцой. Рецептор имеет веретенообразное строение и окружен капсулой. Терминали афферентных нервных миелиновых волокон расположены среди пучков коллагеновых волокон, которые расположены в заполненном жидкостью пространстве. Рецепторы активируются при растяжении сухожилия мышцей, которая сокращается, что приводит к рефлекторному расслаблению этой мышцы.
Схема структуры сухожильного рецептора Гольджи.
В. Рефлекторна функция спинного мозга. Спинальные двигательные рефлексы. В центрах спинного мозга замыкаются рефлекторные дуги двигательных рефлексов: а) тонических, что лежат в основе регуляции тонуса мышц; б) фазических, что обеспечивают перемещение частей тела относительно туловища. Спинальные рефлексы являются сегментарными или межсегментарными. Рефлексы растяжения с мышечных веретен – миотатические:
- возникают при растяжении мышечных веретен в результате растяжения мышц; - информация от мышечных веретен (первичных спиралеподобных нервных окончаний) по афферентным нервным волокнам передается к а-мотонейронам спинного мозга, которые активируются; - активация а-мотонейронов приводит к передаче информации к скелетным мышцам по эфферентним нервным волокнам, следствием этого является их сокращение; - альфа-мотонейроны мышц-антагонистов тормозятся; - рефлекторная дуга миотатичеких рефлексов, которые возникают при растяжении мышечных веретен, является моносинаптической; - при отсутствии нисходящих активирующих влияний двигательных центров головного мозга на а-мотонейроны и гамма-мотонейроны миотатические рефлексы поддерживают мышечный тонус, но он является ослабленным. 2) Рефлексы растяжения с рецепторов сухожилий: - возникают при растяжении сухожилий мышцей, которая сокращается; - информация от рецепторов сухожилий Гольджи передается по афферентным нервным волокнам к тормозным нейронам спинного мозга, которые вызывают постсинаптическое торможение а-мотонейронов мышцы, которая сокращалась, следствием этого является ее расслабление. Рефлекторная дуга рефлекса является полисинаптической.
Схемы рефлекторных дуг: 1) рефлекса растяжения (І) с мышечных веретен (что приводит к сокращению той же мышцы; 2) тормозного рефлекса растяжения с рецепторов сухожилий Гольджи (II), которая имеет тормозной нейрон, который приводит к торможению а-мотонейрона и расслаблению мышцы, которая сокращалась. 3) Флексорные кожно-мышечные рефлексы: - возникают при раздражении рецепторов кожи -информация передается по афферентным волокнам к альфа-мотонейронам спинного мозга, которые активируются; - от альфа-мотонейронов нервные импульсы передаются по эфферентным нервным волокнам к мышцам-флексорам, которые сокращаются, следствием чего есть сгибание конечности и отдаление ее от раздражителя; - рефлекторные дуги кожно-мышечных рефлексов являются полисинаптическими.
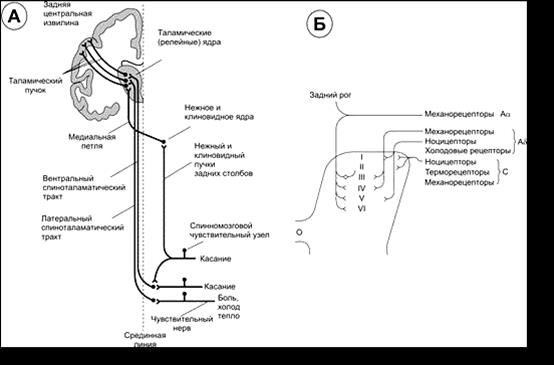
С. Проводниковая функция спинного мозга. Проводящие пути образованы цепями нейронов, которые соединены между собой синапсами, и обеспечивают передачу информации от нейрону к нейрону, от ядра к ядру. Восходящие проводниковые пути
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-12-17; просмотров: 239; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.221.187.121 (0.155 с.) |