Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
На плодовитость и эмбриогенез
Особенность отбора по локусам группоспецифичных антигенов эритроцитов, таких, как «резус» или АВО, заключается в том, что он действует на уровне зиготы или ранних стадий эмбриогенеза. Замечено, что в браках, где мать и отец несовместимы по фактору «резус», число детей с гемолитической болезнью меньше ожидаемого. Так, среди белого населения США Rh-отрицательные люди составляют примерно 16%. В такой популяции частота аллеля d равна 40%, а вероятность развития гемолитической болезни при случайном подборе брачных пар — 9%. На самом деле частота этой болезни составляет 1:150—1:200. Одним из факторов снижения является одновременная несовместимость родителей по локусу АВ0. В этом случае Rh-положительные эритроциты плода, попавшие в организм матери, разрушаются и иммунизации не происходит. Отрицательный отбор действует в большинстве популяций людей по аллелям аномальных гемоглобинов. Его особая жесткость обусловливается тем, что он направлен против гомозигот. Ребенок, умирающий, например, от серповидно-клеточной анемии, является гомозиготным по аллелю S. Каждая такая смерть устраняет из генофонда популяции аллели одного вида. Это приводит к сравнительно быстрому снижению изменчивости по соответствующему локусу. Во многих популяциях людей частота аллелей аномальных гемоглобинов, в том числе и S, не превышает 1%. Высокая частота аллелей таких аномальных гемоглобинов, как S, С, D, Е, в некоторых районах планеты иллюстрирует действие естественного отбора по поддержанию в человеческих популяциях состояния балансированного генетического полиморфизма. Отрицательный отбор в отношении аллеля S перекрывается мощным положительным отбором гетерозигот HbAHbS благодаря высокой жизнеспособности последних в очагах тропической малярии (рис. 12.9).
Рис. 12.9. Совпадение распространения в Старом Свете аллеля серповидно-клеточности (А) и тропической малярии (Б) Исследования в Уганде показали, что количество возбудителей в 1 мл крови зависит от генотипа ребенка и составляет до 10 000 у HbSHbS, до 160 000 у HbAHbS и до 800 000 у НbAHbА. В Северной Греции обследовали 48 семей, в которых наблюдались и серповидно-клеточность, и малярия. Среди братьев и сестер, больных серповидно-клеточной анемией, болело малярией 16 из 25 с генотипами HbAHbA и 1 из 23 с генотипами HbAHbS.
Таким образом, нормальные люди болели в 13 раз чаще, чем индивидуумы с аномальным генотипом. Устранение фактора контротбора приводит к снижению частоты аллеля серповидно-клеточности. Этой причиной, действующей на протяжении уже нескольких столетий наряду с метисацией, объясняют относительно низкую частоту гетерозигот HbAHbS среди североамериканских негров (8—9%) в сравнении с африканскими (около 20%). В приведенных примерах действию отрицательного отбора, снижающего в генофондах некоторых популяций людей концентрацию определенных аллелей, противостоят контротборы, которые поддерживают частоту этих аллелей на достаточно высоком уровне. Результатом наложения многочисленных и разнонаправленных векторов отбора является формирование и поддерживание генофондов популяций в состоянии, обеспечивающем возникновение в каждом поколении генотипов достаточной приспособленности с учетом местных условий. Благодаря социально-экономическим преобразованиям, успехам лечебной и особенно профилактической медицины влияние отбора на генетический состав популяций людей прогрессивно снижается. ГЕНЕТИЧЕСКОЕ РАЗНООБРАЗИЕ В ПОПУЛЯЦИЯХ ЛЮДЕЙ
Человечеству свойствен высокий уровень наследственного разнообразия, что проявляется в многообразии фенотипов. Люди отличаются друг от друга цветом кожных покровов, глаз, волос, формой носа и ушной раковины, рисунком эпидермальных гребней на подушечках пальцев и другими сложными признаками. Выявлены многочисленные варианты отдельных белков, различающиеся по одному или нескольким аминокислотным остаткам и, следовательно, функционально. Белки являются простыми признаками и прямо отражают генетическую конституцию организма. У людей не совпадают группы крови по системам эритроцитарных антигенов «резус», АВ0, MN. Известно более 130 вариантов гемоглобина, более 70 вариантов фермента глюкозо-6-фосфатдегидрогеназы (Г6ФД), который участвует в бескислородном расщеплении глюкозы в эритроцитах. В целом не менее 30% генов, контролирующих у человека синтез ферментных и других белков, имеют несколько аллельных форм. Частота встречаемости разных аллелей одного гена варьирует.
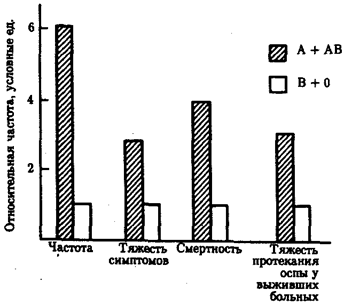
Так, из многих вариантов гемоглобина лишь четыре обнаруживаются в некоторых популяциях в высокой концентрации: HbS (тропическая Африка, Средиземноморье), НЬС (Западная Африка), HbD (Индия), НЬЕ (Юго-Восточная Азия). Концентрация других аллелей гемоглобина повсеместно не превышает, видимо, 0,01—0,0001. Вариабельность распространенности аллелей в популяциях людей зависит от действия элементарных эволюционных факторов. Важная роль принадлежит мутационному процессу, естественному отбору, генетико-автоматическим процессам, миграциям. Мутационный процесс создает новые аллели. И в человеческих популяциях он действует ненаправленно, случайным образом. В силу этого отбор не приводит к выраженному преобладанию концентрации одних аллелей над другими. В достаточно большой популяции, где каждая пара родителей из поколения в поколение дает двух потомков вероятность сохранения новой нейтральной мутации через 15 поколений составляет всего 1/9. Все многообразие вариантов белков, отражающее разнообразие аллелей в генофонде человечества, можно разделить на две группы. К одной из них относятся редкие варианты, встречающиеся повсеместно с частотой менее 1%. Появление их объясняется исключительно мутационным процессом. Вторую группу составляют варианты, обнаруживаемые относительно часто в избранных популяциях. Так, в примере с гемоглобинами к первой группе относятся все варианты, кроме HbS, HbC, HbD и HbE. Длительные различия в концентрации отдельных аллелей между популяциями, сохранение в достаточно высокой концентрации нескольких аллелей в одной популяции зависят от действия естественного отбора или дрейфа генов. К межпопуляционным различиям в концентрации определенных аллелей приводит стабилизирующая форма естественного отбора. Неслучайное распределение по планете аллелей эритроцитарных антигенов АВ0 может быть, например, обусловлено различной выживаемостью лиц, отличающихся по группе крови, в условиях частых эпидемий особо опасных инфекций. Области сравнительно низких частот аллеля I0 и относительно высоких частот аллеля IB в Азии примерно совпадают с очагами чумы. Возбудитель этой инфекции имеет Н-подобный антиген. Это делает людей с группой крови О особенно восприимчивыми к чуме, так как они, имея антиген Н, не способны вырабатывать противочумные антитела в достаточном количестве. Указанному объяснению соответствует факт, что относительно высокие концентрации аллеля I0 обнаруживаются в популяциях аборигенов Австралии и Полинезии, индейцев Америки, которые практически не поражались чумой. Частота заболеваемости «натуральной» оспой, тяжесть симптомов, смертность выше у лиц с группой крови А или АВ в сравнении с лицами, имеющими группу крови 0 или В (рис. 12.10). Объяснение состоит в том, что у людей первых двух групп отсутствуют антитела, частично нейтрализующие оспенный антиген А. Лица с группой крови 0 в среднем имеют возможность прожить дольше, однако для них выше вероятность заболеть язвенной болезнью. В таблице 12.3 указаны некоторые аллели и генотипы, имеющие приспособительное значение в отдельных географических и экологических регионах.
Рис. 12.10. Относительная частота и показатели тяжести заболевания «натуральной» оспой у лиц с разными группами крови АВО
Таблица 12.3. Примеры аллелей, имеющих адаптивное значение
Вместе с тем для популяций из одного географического района, но изолированных в репродуктивном отношении, причиной различий в концентрации аллелей АВО мог быть дрейф генов. Так, частота группы крови А достигает у индейцев племени черноногих 80%, а у индейцев из штата Юта — 2%. В основе стойкого сохранения в популяции людей одновременно нескольких аллелей одного гена лежит, как правило, отбор в пользу гетерозигот, который ведет к состоянию балансированного полиморфизма. Классическим примером такой ситуации является распространение аллелей гемоглобинов S, С, и Е в очагах тропической малярии. Выше приведены примеры полиморфизма по конкретным локусам, который объясняется действием известного фактора отбора. В естественных условиях в силу воздействия на фенотипы организмов комплекса факторов отбор осуществляется по многим направлениям. В результате формируются генофонды, сбалансированные по набору и частотам аллелей, обеспечивающие в данных условиях достаточную выживаемость популяций. Это справедливо и для популяций людей. Так, люди с группой крови 0 более восприимчивы к чуме, чем люди с группой В. Туберкулез легких у них лечится с большим трудом, чем у лиц с группой крови А. Вместе с тем лечение больных сифилисом людей с группой крови 0 быстрее вызывает переход болезни в неактивную стадию. Для лиц с группой крови 0 вероятность заболеть раком желудка, раком шейки матки, ревматизмом, ишемической болезнью сердца, холециститом, жёлчно-каменной болезнью примерно на 20% ниже, чем для лиц с группой А.
Генетический полиморфизм по многим локусам мог быть унаследован людьми от предков на досапиентной стадии развития. Полиморфизм по таким системам групп крови, как АВ0 и Rh, обнаружен у человекообразных обезьян. Факторы отбора, действие которых создавало современную картину распределения аллелей в популяции людей, для подавляющего большинства локусов точно не установлены. Примеры, рассмотренные выше, указывают на их экологическую природу. Учитывая слабую техническую оснащенность, плохие экономические и гигиенические условия жизни основной массы населения планеты на протяжении значительной части истории человечества, можно представить, какую большую роль играли возбудители особо опасных инфекций, паразитарных заболеваний, туберкулеза. В этих условиях наследственный полиморфизм способствовал расселению людей, обусловливая удовлетворительную жизнеспособность в разных экологических ситуациях. Определенный вклад в наблюдаемое распределение аллелей внесли массовые миграции населения и сопутствующая им метизация. В период до Великих географических открытий и начала колониальной экспансии смешение больших контингентов людей разной расовой принадлежности имело место в Восточной Африке, Индии, Средней Азии, Западной Сибири, Алтае-Саянском нагорье, Индокитае. Впоследствии это наблюдалось в Южной и Центральной Америке. Генетический полиморфизм является основой межпопуляционной и внутрипопуляционной изменчивости людей. Изменчивость проявляется в неравномерном распределении по планете некоторых заболеваний, тяжести их протекания в разных человеческих популяциях, разной степени предрасположенности людей к определенным болезням, индивидуальных особенностях развития патологических процессов, различиях в реакции на лечебное воздействие. Наследственное разнообразие долго было препятствием успешному переливанию крови. В настоящее время оно же создает большие трудности в решении проблемы пересадок тканей и органов. ГЕНЕТИЧЕСКИЙ ГРУЗ В ПОПУЛЯЦИЯХ ЛЮДЕЙ
Так же как и в популяциях других организмов, наследственное разнообразие снижает реальную приспособленность популяций людей. Бремя генетического груза человечества можно оценить, введя понятие летальных эквивалентов. Считают, что число их в пересчете на гамету колеблется от 1,5 до 2,5 или от 3 до 5 на зиготу. Это означает, что то количество неблагоприятных аллелей, которое имеется в генотипе каждого человека, по своему суммарному вредному действию эквивалентно действию 3—5 рецессивных аллелей, приводящих в гомозиготном состоянии к смерти индивидуума до наступления репродуктивного возраста.
При наличии неблагоприятных аллелей и их сочетаний примерно половина зигот, образующихся в каждом поколении людей, в биологическом плане несостоятельна. Такие зиготы не участвуют в передаче генов следующему поколению. Около 15% зачатых организмов гибнет до рождения, 3 — при рождении, 2 — непосредственно после рождения, 3 — умирает, не достигнув половой зрелости, 20 — не вступают в брак, 10% браков бездетны. Неблагоприятные последствия генетического груза в виде рецессивных аллелей, если они не приводят к гибели организма, проявляются в снижении ряда важных показателей состояния индивидуума, в частности его умственных способностей. Исследования, проведенные на популяции арабов в Израиле, для которой характерна высокая частота близкородственных браков (34% между двоюродными и 4% между дважды двоюродными сибсами), показали снижение умственных способностей у детей от таких браков. Исторические перспективы человека в силу его социальной сущности не связаны с генетической информацией, накопленной видом Homo sapiens в ходе эволюции. Тем не менее человечество продолжает «оплачивать» эти перспективы, теряя в каждом поколении часть своих членов из-за их генетической несостоятельности. ГЛАВА 13
|
||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 218; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.16.147.124 (0.02 с.) |