Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Физиологии возбудимых тканейСтр 1 из 7Следующая ⇒
В.А. Пестряев КОНСПЕКТ ЛЕКЦИЙ ПО ФИЗИОЛОГИИ ВОЗБУДИМЫХ ТКАНЕЙ ЕКАТЕРИНБУРГ
Оглавление Список используемых сокращений…………………………………………………...
2.1. Потенциал покоя……………………………………………………………………. 2.2. Потенциал действия……………………………………………………………….…
4.1. Механизмы потенциала покоя……………………………………………………… 4.2. Механизмы потенциал действия……………………………………………………
6.1. Закон анатомической и физиологической непрерывности нервного волокна…... 6.2. Закон двустороннего проведения возбуждения…………………………………… 6.3. Закон изолированного проведения возбуждения в нервных стволах…………….
7.1. Закон силы……………………………………………………………………………. 7.2. Зависимость пороговой силы от длительности стимула………………………….. 7.3. Зависимость порога от крутизны нарастания раздражителя……………………… 7.4. Закон «все или ничего»……………………………………………………………… 7.5. Полярный закон раздражения ……………………………………………………… 7.6. Лабильность …………………………………………………………………………. 8. Межклеточная передача возбуждения. Синапс………………………………….…… 8.1.Химический синапс…………………………………………………………………… 8.2.Электрическая передача…………..………………………………………………….. 8.2.1.Электрический синапс…………………………………………………………….. 8.2.2.Эфаптическая передача…………………………………………………………… 9. Возникновение потенциала действия в афферентных нейронах. Рецепторный и генераторный потенциалы………………………………………….. 10. Возникновение потенциала действия в эфферентных нейронах. Механизмы суммации постсинаптических потенциалов………………………….. 11. Виды торможения в ЦНС…………………………………………………………….
12. Скелетные мышцы…………………………….…………..……………………….… 13 Сердечная мышца……………………………………………………………………. 14. Гладкие мышцы……………………………………………………………………… 15. Гландулоциты……………………………………………………………………….. Список используемых сокращений.
АР - адренорецепторы АРП - абсолютный рефрактерный период АТФ - аденозинтрифосфат АТФ-аза - аденозинтрифосфатаза АЦХ - ацетилхолин АЦХЭ - ацетилхолинэстераза ВПСП - возбуждающий постсинаптический потенциал ГАМК - гамма-аминомасляная кислота ГМК - гладкомышечные клетки КУД - критический уровень деполяризации МДД - медленная диастолическая деполяризация МП - мембранный потенциал МПП - мембранный потенциал покоя МПКП - миниатюрный потенциал концевой пластинки МПП - мембранный потенциал покоя М-ХР - мускариновые холинорецепторы НА - норадреналин Н-ХР - никотиновые холинорецепторы ПД - потенциал действия ПКП - потенциал концевой пластинки ПП - потенциал покоя ПреСМ - пресинаптическая мембрана ПСМ - постсинаптическая мембрана ПСП - постсинаптический потенциал СДП - спонтанная диастолическая деполяризация ТПСП - тормозной постсинаптический потенциал ЦНС - центральная нервная система
ПОТЕНЦИАЛ ПОКОЯ И ПОТЕНЦИАЛ ДЕЙСТВИЯ 2.1. Потенциал покоя. Микроэлектродная техника (внутриклеточная регистрация биопотенциалов). Микроэлектрод – стеклянная микропипетка, заполненная раствором электролита. Диаметр кончика менее 0,5 мкм позволяет ввести электрод внутрь клетки, не нарушая ее функции. Второй электрод (электрод сравнения) – в питающий раствор с исследуемой тканью. Потенциал внеклеточной среды принимается равным нулю. Электроды соединяются с согласующим устройством, потом с усилителем постоянного тока. В качестве регистратора используется осциллограф. В момент прокола мембраны клетки микроэлектродом на экране осциллографа происходит резкое смещение нулевого уровня книзу (рис 3). Наблюдается поляризация мембраны - внутренняя сторона мембраны заряжена отрицательно относительно внешней. Это же касается заряда внутреннего содержимого клетки относительно внешней среды. Перемещение кончика микроэлектрода внутри клетки не приводит к изменению измеряемой разности потенциалов, если электрод не повредил клетку. Зарегистрированная разность потенциалов получила название потенциала покоя (ПП) или мембранного потенциала покоя (МПП).
Обычно величина МПП колеблется от – 70 до – 95 мВ. Смещение мембранного потенциала кверху, т.е. уменьшение значения мембранного потенциала по модулю (уменьшение поляризации) называется уменьшением мембранного потенциала или деполяризацией; смещение книзу, т.е. увеличение по модулю значения мембранного потенциала (увеличение поляризации), называется увеличением мембранного потенциала или гиперполяризацией (рис. 4). Потенциал действия. При неизменном функциональном состоянии клетки величина мембранного потенциала не изменяется. Поддержание постоянной его величины обеспечивается нормальным протеканием клеточного метаболизма.
Рис. 3. Внутриклеточная регистрация мембранного потенциала. А – схема установки для регистрации; Б – момент введения микроэлектрода в клетку. 1 – стеклянный микроэлектрод; 2 – электрод сравнения; 3 – усилитель; 4 – регистратор.
Рис. 4. Изменения поляризации (потенциала) мембраны.
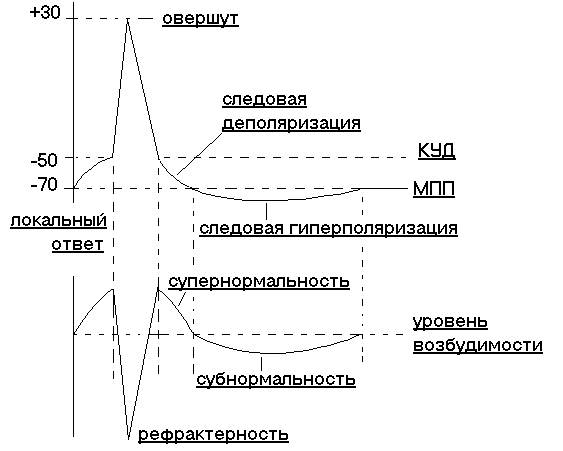
При нанесении на клетку, в которой находится микроэлектрод, допороговых стимулов, можно зарегистрировать уменьшение мембранного потенциала (деполяризацию), которое обратимо (быстро проходит) и зависит от силы стимула, но до определенного уровня. Ответы клетки при действии на нее допороговых раздражений могут суммироваться. При деполяризации до определенного уровня (обычно – это смещение мембранного потенциала на 20-30% от величины МПП), называемого критический уровень деполяризации (КУД), возникает резкое колебание мембранного потенциала (рис 5), получившее название потенциала действия (ПД) или спайка или пик – потенциала. И как бы мы дальше не увеличивали силу раздражения, амплитуда потенциала действия уже не изменится (закон “все или ничего”). Все изменения мембранного потенциала до КУД отображают местный процесс возбуждения, нераспространяющееся возбуждение или локальный ответ. В ПД различают пик и следовые потенциалы. Восходящая часть пика – деполяризация, нисходящая – реполяризация. Овершут – перезарядка мембраны или перескок – основная причина распространения возбуждения. Именно эти овершуты, перескоки ПД и регистрировал в своих экспериментах Эмиль Дюбуа-Реймон. ПД – это всегда распространяющееся возбуждение. Следовые потенциалы: отрицательный следовый потенциал (следовая деполяризация); положительный следовый потенциал (следовая гиперполяризация). Амплитуда потенциала действия: нервные клетки 110 –100 мВ; скелетные и сердечные мышцы 110 – 120 мВ. Продолжительность ПД нервных клеток 1 –2 мс. Фазовые изменения возбудимости при генерации ПД (рис. 5). Мерило возбудимости – порог раздражения. При местном, локальном, возбуждении возбудимость увеличивается. Т.е. когда мембранный потенциал достигает КУД, возбудимость повышена. ПД сопровождается многофазными изменениями возбудимости: Период абсолютной рефрактерности (АРП) соответствует фазе деполяризации потенциала действия, пику и началу фазы реполяризации, возбудимость снижена вплоть до полного отсутствия во время пика.
Период относительной рефрактерности соответствует оставшейся части фазы реполяризации, возбудимость постепенно восстанавливается к исходному уровню. Супернормальный период соответствует фазе следовой деполяризации потенциала действия (отрицательный следовый потенциал), возбудимость повышена. Субнормальный период соответствует фазе следовой гиперполяризации потенциала действия (положительный следовый потенциал), возбудимость снижена. Если потенциал покоя присущ всем живым клеткам без исключения, то потенциал действия генерируется только возбудимыми клетками, является электрофизиологическим показателем возникновения и распространения процесса возбуждения по мембранам нервных и мышечных клеток.
Рис. 5. ПД и изменения возбудимости во время ПД.
Потенциал покоя. Теория, объясняющая происхождение мембранного потенциала, базируется на двух основных положениях: 1. Мембрана избирательно проницаема для различных ионов. В состоянии покоя мембрана проницаема для катионов и практически непроницаема для анионов. В покое проницаемость для ионов калия гораздо выше, чем для ионов натрия. 2. В мембране имеется биохимический механизм, получивший название “натриевого насоса” - натрий-калиевая АТФ-аза, обеспечивающий активный транспорт ионов калия внутрь клетки, а ионов натрия наружу. В состоянии покоя в цитоплазме клетки ионов натрия в 10 раз меньше, чем снаружи, а ионов калия в 50 раз больше, чем снаружи. Это состояние поддерживается работой насоса. Работает насос против градиента концентрации с затратой энергии. В покое мембрана имеет более высокую проницаемость для ионов калия, чем для ионов натрия и тем более анионов, внутри клетки. По закону осмоса, несмотря на работу насоса, калий стремится выйти из клетки, а анионы не могут последовать за ним, что приводит к разделению зарядов и появлению на мембране потенциала, отрицательного внутри и положительного снаружи. Величина мембранного потенциала зависит от концентрации ионов калия внутри клетки и снаружи и может быть вычислена на основе законов физической химии (уравнение Нернста и уравнение Гольдмана – Ходжкина – Катца). Работа натриевого насоса при гидролизе одной молекулы АТФ сопровождается выведением из клетки трех ионов натрия и поглощением двух ионов калия. Поскольку при этом перенос зарядов не скомпенсирован, то в результате функционирования АТФ – азы на мембране клетки разность потенциалов суммируется.
Кроме того, в создании отрицательного заряда принимают участие и белковые молекулы протоплазмы клетки. Потенциал действия. Возникновение ПД связано с изменением проницаемости клеточной мембраны при ее возбуждении. При деполяризации мембраны до некоторого критического уровня (КУД) открываются натриевые каналы и ионы натрия по градиенту концентрации, без затрат энергии устремляются внутрь клетки, обусловливая фазу деполяризации потенциала действия. Этот лавинообразный поток ионов натрия внутрь клетки продолжается до момента перезарядки мембраны. Во время овершута (перескока) наступает резкое снижение проницаемости для натрия (натриевая инактивация), но резко увеличивается проницаемость мембраны для ионов калия, которые по градиенту концентрации без затрат энергии, выходят из клетки, компенсируя вошедшие положительно заряженные ионы натрия и обусловливая возвращение мембранного потенциала на исходный уровень (фаза реполяризации). Таким образом, по заряду (потенциалу) клетка вернулась на исходный уровень, а ионный состав ее нарушен. Внутри увеличилось количество ионов натрия, а снаружи увеличилось количество ионов калия. Это именно та ситуация, когда натриевый насос работает наиболее активно, восстанавливая ионное равновесие (точнее ионное неравновесие) клетки. Возникновение потенциала действия связано в основном с движением ионов натрия внутрь. Поэтому ПД считают “натриевым потенциалом”, в отличие от потенциала покоя, который считается в основном “калиевым”.
Закон силы. Возникновение распространяющегося возбуждения (ПД) возможно при условии, когда действующий на клетку раздражитель имеет некоторую минимальную (пороговую силу), иначе говоря, когда сила раздражителя соответствует порогу раздражения. Порог – это та наименьшая величина раздражителя, которая действуя на клетку какое-то определенное время, способна вызвать максимальное возбуждение; - это та наименьшая величина раздражителя, при действии которой потенциал покоя может сместиться до КУД мембраны, при котором активируется перенос ионов натрия внутрь клетки.
Закон “ все или ничего”. Установлен Боудичем в 1871 г. на мышце сердца. При подпороговой силе раздражения мышца сердца не сокращается, а при пороговой силе раздражения – сокращение максимально. При дальнейшем увеличении силы раздражения амплитуда сокращений не увеличивается. Со временем была установлена универсальность этого закона по отношению ко всем возбудимым тканям. Однако исследования с использованием микроэлектродной техники выявили и некоторое формальное несоответствие: подпороговое раздражение вызывает местное, нераспространяющееся возбуждение, следовательно, нельзя говорить, что допороговое раздражение не дает ничего.
Процесс развития возбуждения подчиняется этому закону с КУД, когда запускается лавинообразное поступление ионов натрия в клетку.
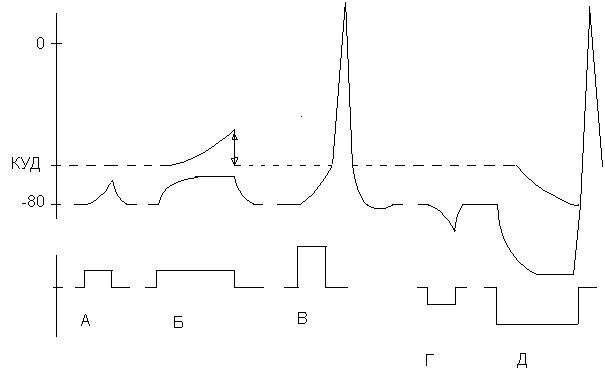
7.5. Полярный закон раздражения (закон Пфлюгера). Законы Пфлюгера (1859) основываются на изменениях мембранного потенциала при действии на возбудимые ткани постоянного электрического тока (рис. 10).
Рис. 10. Действие электрического тока на возбудимые ткани. А – изменение МП под катодом при кратковременном пропускании тока; Б – при длительном пропускании тока; В – возникновение ПД при пороговом значении тока; Г – изменение МП под анодом при кратковременном пропускании тока; Д – изменение МП и КУД при длительном действии сильного анодного тока – анодно-размыкательное возбуждение.
1. Постоянный ток проявляет свое раздражающее действие только в момент замыкания и размыкания цепи. 2. При замыкании цепи постоянного тока возбуждение возникает под катодом; при размыкании под анодом.
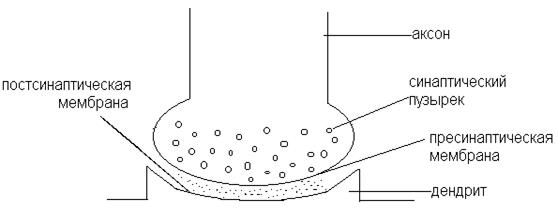
СИНАПС Синапсами называют контакты, которые устанавливают нейроны. Синапс – это специализированная структура, обеспечивающая передачу нервного импульса с аксона на другую клетку. Существуют классификации синапсов по местоположению, по характеру действия, по способу передачи сигнала. По местоположению: нервно-мышечные (мионейрональные), нейросекреторные, нейронейрональные (межнейронные). Нейрональные: аксосоматические, аксодендритические, аксоаксональные. По характеру действия: возбуждающие и тормозящие. По способу передачи сигнала: электрические, химические, смешанные. Химические синапсы. У млекопитающих и человека в НС чаще встречаются химические синапсы (рис. 11), которые состоят из пресинаптической части (окончание нейрона, передающего сигнал), синаптической щели (непосредственное продолжение межклеточного пространства) и постсинаптической части (структура воспринимающей клетки). При поступлении возбуждения (ПД) к окончанию аксона, в нем высвобождается химическое вещество (медиатор), которое вызывает возбуждение или торможение на мембране иннервируемой клетки. В синапсе возбуждение всегда передается от пресинаптического (аксонного) участка к постсинаптической области соседней клетки (рис. 11).
Рис. 11. Химический синапс.
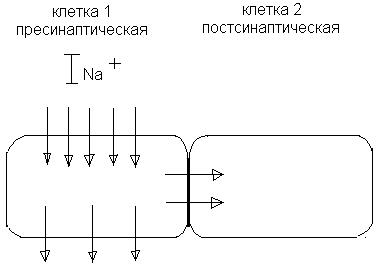
Электрическая передача. Электрические синапсы. Встречаются редко. ПД вызывает возбуждение в соседней клетке или торможение без химического посредника. Концептуально были предсказаны еще до открытия химических синапсов (назывались эфапсами). Между 1930 – 1950 г.г., когда концепция химической синаптической передачи была уже общепризнанной, выяснилось, что межклеточная передача возбуждения может осуществляться и электрическим способом. Принцип: 2 соседние клетки прилегают тесно друг к другу (рис. 12). Сопротивление электрическому току соприкасающихся мембран должно быть сравнимо с сопротивлением остальной, внесинаптической области мембраны. При возбуждении клетки 1 натриевый ток входит в нее через открытые натриевые каналы и выходит через пока не возбужденные участки мембраны. Часть тока входит через участок мембранного контакта в клетке 2, вызывая ее деполяризацию. Уровень деполяризации здесь примерно в 10 раз ниже, чем в клетке 1, однако выше порога генерирования ПД в клетке 2. Часто такая деполяризация подпороговая, а клетка 2 возбуждается в результате суммации синаптических потенциалов.
Рис. 12. Электрический синапс.
Ионы, переносящие электрические токи не могут проходить через липидные мембраны. Для их транспорта в «мембранных контактах» между электрически сопряженными клетками необходимы канальные белки. Такие межклеточные связи называются нексусами, или «щелевыми контактами». Щелевые контакты соединяют группы синхронно функционирующих клеток. В тканях не относящихся к НС, клетки также бывают соединены щелевыми контактами. В миокарде и гладкой мускулатуре эти контакты создают функциональный синцитий. Возбуждение здесь переходит от одной клетки к другой без заметной паузы или снижения амплитуды ПД на границе. Для таких органов важна регулируемость щелевых контактов. Их каналы закрываются при снижении рН или повышении концентрации Са2+. Это происходит в случае повреждения клеток или глубокого нарушения обмена. За счет такого механизма пораженные места изолируются от остальной части функционального синцития, и распространение патологии ограничивается (например, при инфаркте миокарда). Кроме этих возбудимых тканей существует и много других, где клетки также соединены щелевыми контактами (все эпителии, печень). В принципе такая связь присуща любой клетке на ранних стадиях эмбрионального развития, когда все клетки соединены между собой щелевыми контактами и сохраняют их до стадии дифференцировки органов. Щелевой контакт – наиболее распространеный тип электрического синапса. Однако существуют и другие. Например, электрическим путем может передаваться и торможение. В этом случае ПД особым образом расположенных пресинаптических волокон генерируют во внеклеточном пространстве вокруг постсинаптического аксона местный положительный потенциал такой амплитуды, что его деполяризация не может достичь порогового уровня, и проведение по нему ПД блокируется. Учитывая широкое распространение щелевых контактов, кажется удивительным, почему в нервной системе они не используются для синаптической передачи повсеместно. Видимо, сложнее организованные химические синапсы обеспечивают настолько более высокую специфичность и регулируемость межклеточной коммуникации, что в значительной степени вытеснили электрические.
Эфаптическая передача. При некоторых заболеваниях аксоны теряют свою миелиновую оболочку, становясь демиелинизированными. Демиелинизированные аксоны могут вступать в аномальные взаимодействия, когда импульсы, проходящие по группам нервных волокон, индуцируют возбуждение других параллельно идущих аксонов. Это называется эфаптической передачей. Когда такие аномальные ПД генерируются в сенсорных нервных волокнах, появляются аномальные ощущения, парестезии. Если они связаны с ноцицептивными (болевыми) волокнами, то возникают такие неприятные синдромы, как невралгия, каузалгия, невромные боли. Межаксонные помехи могут быть следствием не только недостаточной изоляции (миелиновыми оболочками), но и повышенной возбудимости аксонов.
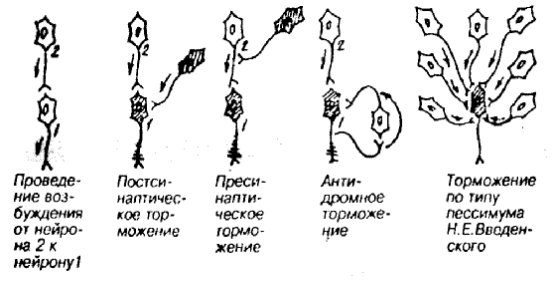
ВИДЫ ТОРМОЖЕНИЯ В ЦНС Различают несколько видов торможения: постсинаптическое, пресинаптическое, пессимальное (рис. 14). Постсинаптическое торможение – развивается в ПСМ аксосоматичесих и аксодендритеческих синапсов под влиянием тормозных нейронов, у которых из концевых разветвлений аксонных отростков в синаптическую щель высвобождается тормозной медиатор (например, ГАМК или глицин). Медиатор вызывает в ПСМ эффект гиперполяризации в виде ТПСП, пространственно-временная суммация которых приводит к урежению или прекращению генерации ПД. Пресинаптическое торможение – развертывается в аксоаксональных синапсах, блокируя распространение возбуждения по аксону. Процесс торможения здесь протекает по типу катодической депрессии: в области контакта выделяется медиатор (ГАМК), который вызывает стойкую деполяризацию, что нарушает проведение волны возбуждения через этот участок. Является наиболее избирательным видом торможения. Пессимальное торможение – вид торможения центральных нейронов. Наступает при высокой частоте раздражения. В первый момент возникает высокая частота ответного возбуждения, однако через некоторое время стимулируемый в таком режиме нейрон переходит в состояние торможения. Возвратное (антидромное) торможение – угнетение активности нейрона вызывается возвратной коллатералью аксона, заканчивающейся на тормозном нейроне, аксон которого оказывает тормозное действие.
Рис. 14. Виды внутрицентрального торможения.
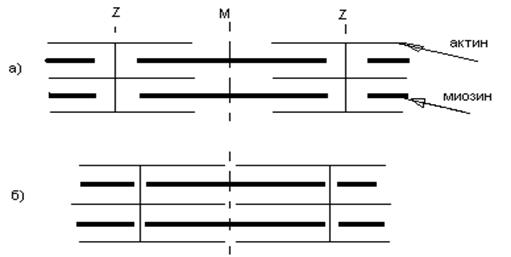
СКЕЛЕТНЫЕ МЫШЦЫ Скелетные мышцыобеспечивают перемещение составных элементов скелета. Имеют поперечную исчерченность (поперечнополосатые). Свойства скелетной мышцы: 1) возбудимость; 2) проводимость (способность проводить ПД вдоль мышечного волокна и вглубь его по Т-системе поперечных трубочек, служащих связующим звеном между поверхностной мембраной и сократительным аппаратом); 3) сократимость (способность укорачиваться или развивать напряжение при возбуждении); 4) эластичность (способность развивать напряжение при растягивании). Скелетные мышцы имеют 2 типа волокон: интрафузальные и экстрафузальные. Интрафузальное волокно находится внутри мышечного веретена (специализированного мышечного рецептора), располагающегося в толще скелетной мышцы. Оно необходимо для регуляции чувствительности рецептора и управляется специальными мотонейронами спинного мозга – гамма-мотонейронами. Мышечные волокна не входящие в состав мышечного веретена, называются экстрафузальными. Суммированные сокращения. В зависимости от частоты раздражения меняется характер сокращения. Если очередной стимул (или его действие) попадает в фазу расслабления, мышца не успевает расслабиться, возникает дополнительное сокращение, развивается длительное напряжение - зубчатый тетанус. При более высокой частоте (т.е. с еще меньшим интервалом между раздражителями), когда каждый очередной стимул попадает в фазу укорочения мышцы, происходит продолжительная активация сократительной системы, развивается мощное длительное сокращение, которое называется гладким тетанусом. Расслабление возникает при утомлении. Амплитуда гладкого тетануса зависит от частоты раздражения. Если каждый последующий стимул (раздражитель) попадает в фазу экзальтации (повышенной возбудимости), ответ мышцы будет достаточно большим, если же импульсы попадают в период сниженной возбудимости (относительная рефрактерная фаза), то ответ мышцы будет намного меньше. Напр. 30 Гц – 10 мм, 50 Гц – 15 мм, 200 Гц – 3 мм. Такая зависимость амплитуды ответа мышцы от частоты получила название оптимума и пессимума частоты раздражения. Альфа-мотонейрон может посылать к мышце серию импульсов, например, 20 имп/с, 40 имп/с, 50 имп/с. Все наши сокращения в ответ на импульсную стимуляцию частотного характера являются тетаническими. Структура саркомера. С помощью светового микроскопа в миофибрилле обнаружены правильно чередующиеся поперечные светлые и темные полосы (исчерченность), обусловленные особой регулярной организацией или расположением нитей актина и миозина в саркомерах. В середине такого саркомера располагается пучок толстых нитей миозина. Исчерченность обусловлена правильной организацией актина и миозина. В середине – толстые нити миозина, нити актина жестко закреплены в Z- мембранах по типу щетина в щетках. Именно Z-мембраны ограничивают отдельный саркомер скелетной мышцы. Более темные участки А-диски (анизотропные) обладают двойным лучепреломлением. В их центре видна более светлая полоска (Н-зона). По обе стороны А-диска светлые изотропные полоски - I-диски, образованные нитями актина. В центре Н-полоски обнаружена М-линия – структура, которая удерживает нити миозина. Укорочение саркомеров. Мышца укорачивается в результате сокращения множества саркомеров, соединенных последовательно. При укорочении тонкие актиновые нити скользят вдоль толстых миозиновых и двигаются к середине саркомера (рис. 15). Во время скольжения длина актиновых и миозиновых нитей не меняется; при наблюдении в световой микроскоп не изменяется ширина А-диска, тогда как I-диски и Н-зона становятся более узкими.
Рис. 15. Скольжение нитей актина и миозина при сокращении саркомера. а) – мышца в покое; б) – при сокращении. Работа поперечных мостиков. Миозиновые нити имеют поперечные выступы, которые представляют собой субфрагменты миозина – тяжелый меромиозин, в котором различают шейку и головку. Эти ферменты обладают АТФ-азной активностью (способностью расщеплять АТФ), они отходят биполярно. Во время сокращения каждый поперечный мостик может связываться с актиновой нитью. В момент взаимодействия головки с актиновой нитью развивается усилие, которое сопровождается поворотом головки на 45º, т. е. она действует как рычаг, приводя в движение актиновую нить. Биполярное расположение головок в обеих половинах саркомеров приводит к скольжению актиновых нитей в правой и левой половинах саркомера. В момент соединения поперечного мостика с актиновой нитью происходит активация АТФ-азы этого мостика и затем расщепление АТФ. Предполагают, что энергия расщепления АТФ необходима для разделения актина и миозина. Расщепление – обязательное условие, которое обеспечивает следующий цикл взаимодействия актина и поперечных мостиков. Таким образом, происходит ритмическое отсоединение и присоединение головок миозина к актиновым нитям (сходство с группой людей, которая тянет длинную веревку). Несмотря на ритмичную смену прикрепления и отсоединения поперечных мостиков с частотой от 5 до 50 Гц, сила, развиваемая мышцей в физиологических условиях не колеблется, так как гребковые движения поперечных мостиков происходят асинхронно. В случае уменьшения концентрации АТФ цикличность может нарушаться, а существенное снижение концентрации АТФ может привести к устойчивому прикреплению мостиков к актину. Этим объясняется состояние трупного окоченения (расслабление будет возможно в результате аутолиза). Электромеханическое сопряжение: 1. Генерация ПД. (Стимуляция приводит к деполяризации сарколеммы.) 2. Распространение ПД по Т-системе. (Деполяризация Т-системы и саркоплазматического ретикулума.) 3. Электрическая стимуляция зоны контакта Т-системы и саркоплазматического ретикулума, активация ферментов, образование инозитолтрифосфата, повышение внутриклеточной концентрации ионов кальция. (Выход ионов кальция из саркоплазматического ретикулума.) Сокращение: 4. Образуется комплекс кальций + тропонин. Комплекс кальций + тропонин снимает блокаду актина тропомиозином (освобождение активных центров на актиновых филаментах), а также снимает блокаду тропонином I АТФ-азной активности миозина. 5. Взаимодействие миозиновой головки с актином, вращение головки и развитие эластической тяги. 6. Скольжение нитей актина и миозина относительно друг друга, уменьшение размера саркомера, развитие напряжения или укорочение мышечного волокна. Расслабление: 7. Кальций отделяется от комплекса с тропонином. 8. Кальций диффундирует от тонких филаментов в саркоплазматический ретикулюм. 9. Тропомиозин возвращается на блокирующее место. 10. Тропонин I блокирует АТФ-азную активность миозина. 11. Поперечные актомиозиновые мостики разрываются и нити смещается друг относительно друга. В головках вновь накапливается АТФ. Таким образом, в механизме сокращения скелетных мышц сократительными белками являются - актин и миозин, а регуляторными - тропонин и тропомиозин.
Контрактура -в условиях целостного организма возникает при патологии и проявляется в длительном, слитном сокращении мышцы, которое не управляется корой (волей человека). Природа контрактур различна. В экспериментальных условиях ее легко получить путем воздействия на мышцы. Гиперкалиевый раствор: вызывает длительную деполяризацию мембраны, что приводит к достаточно длительной активности мышцы (калиевая контрактура). Кофеиновая контрактура: длительное сокращение, которое держится в течении всего времени, пока в растворе содержится кофеин. Является следствием высвобождения ионов кальция из саркоплазматического ретикулума.
СЕРДЕЧНАЯ МЫШЦА Сердечная мышца (миокард) относится к возбудимым тканям и имеет поперечную исчерченность. Однако в области Z-линий имеются участки слияния (переплетения) волокон (в этих участках образуются вставочные диски). Благодаря этой особенности сердечная мышца представляет собой сеть волокон. Т-система кардиомиоцитов локализована в области Z-линий (а не на месте слияния А и I-дисков, как в скелетной). В ответ на раздражение сердечная мышца сокращается в соответствии с законом «все или ничего», т.е. либо с максимальной силой, либо не сокращается вовсе. У скелетной мышцы закон «всё или ничего» выполняется применительно к отдельному мышечному волокну, а не к мышце в целом, т.к. отдельные волокна имеют разную возбудимость. Свойства сердечной мышцы: 1) возбудимость; 2) проводимость; 3) сократимость; 4) автоматия. Автоматия – это значит, что возбуждение возникает в сердце периодически, под влиянием процессов, протекающих в нём самом. Однако способностью к автоматии обладают лишь определённые участки миокарда, состоящие из атипичной мышечной ткани, бедной миофибриллами. Все мышечные клетки сердца можно разделить на два больших класса: 1) миокардиоциты – осуществляют сокращение в ответ на приходящий ПД; 2) миоциты – входят в состав узлов автоматии и проводящей системы. Для последних способность к сокращению выражена слабо, главная их функция – генерация автоматического ПД и быстрое проведение возбуждения по сердцу. Мембранный потенциал атипичных волокон не держится на стационарном уровне, происходит медленный сдвиг мембранного потенциала в сторону КУД – фаза спонтанной (или медленной) диастолической деполяризации (СДД или МДД). МДД – внутриклеточный механизм спонтанного возбуждения клеток, лежит в основе автоматии. При достижении КУД начинается генерация ПД, затем снова МДД провоцирует появление очередного потенциала действия и т.д.. Причины фазы МДД: 1) Снижение активности натрий-калиевой АТФ-азы; 2) Низкая проницаемость мембраны для ионов калия; 3) Высокая проницаемость мембраны для ионов натрия. Проводящая система. В норме возбуждение возникает в синоатриальном(синусно-предсердном ) узле - в стенке правого предсердия у места впадения в него верхней полой вены. В атипичных клетках синоатриального узла наиболее высокая скорость МДД. Частота генерации ПД 60-80 в мин. Он навязывает ритм деятельности сердцу – является пейсмекером 1-го порядка. От синоатриального узла возбуждение распространяется (скорость 0,8 – 0,9 м/с) по волокнам правого и левого предсердия, запуская процесс их сокращений. От предсердий возбуждение достигает перегородки между предсердиями и желудочками и попадает в атриовентрикулярный (предсердно-желудочковый) узел. В сердце теплокровных животных существуют специальные проводящие пути между синоатриальным и атриовентрикулярным узлами, а также между правым и левым предсердиями. В атриовентрикулярном узле значительно замедляется скорость проведения возбуждения (0,02 – 0,05 м/с). Эта атриовентрикулярная задержка проведения необходима для того, чтобы отставить во времени процесс возбуждения в предсердиях и желудочках: камеры предсердий и желудочков работают поочередно. От атриовентрикулярного узла возбуждение переходит на пучок Гиса. Благодаря атриовентрикулярной задержке возбуждение доходит до пучка Гиса тогда, когда предсердия успевают сократиться. Пучок Гиса прободает предсердно-желудочковую перегородку и делится на правую и левую ножки. Ножки следуют в межжелудочковой перегородке, а в области верхушки сердца загибаются вверх и переходят в сеть сердечных проводящих миоцитов (волокон Пуркинье), погружённых в рабочий (сократительный) миокард желудочков. Скорость распространения возбуждения в пучке Гиса (4,5 – 5 м/с) и проводящих миоцитах в 5 раз больше скорости распространения по рабочему миокарду. Благодаря этому, миокардиоциты желудочков вовлекаются в сокращение почти одновременно. Существует градиент автоматии, выражающийся в убывающей способности к автоматии (из-за понижения скорости МДД) различных участков проводящей системы, по мере удаления от синоатриального узла. При повреждении синоатриального узла роль водителя ритма – пейсмекера 2-го порядка берёт на себя атриовентрикулярный узел (частота разрядов 30 – 40 в мин.). Вентрикулярная проводящая система – пучок Гиса, волокна Пуркинье – пейсмекеры 3-го порядка (частота 15-20 в мин.). Проводящая система обеспечивает: 1) ритмическую генерацию ПД; 2) координацию сокращений предсердий и желудочков; 3) синхронное вовлечение в процесс сокращения миокардиоцитов желудочков. Отличительная особенность проведения возбуждения в миокарде – большое количество межклеточных контактов в виде специальных структурных образований – вставочных дисков. Межклеточные контакты – нексусы благодаря низкому сопротивлению электрическому току обеспечивают быстрый переход возбуждения с одной клетки на другую. Благодаря нексусам мышечная ткань предсердий и желудочков ведёт себя как функциональный синцитий: возбуждение, возникнув в каком-либо из отделов, охватывает все без исключения невозбуждённые волокна. Сокращение сердца, как и скелетных мышц, запускается ПД. Однако, временные соотношения этих процессов в этих двух типах мышц различны. Длительность ПД скелетных мышц составляет несколько мс, а сокращение начинается тогда, когда возбуждение уже почти заканчивается. В миокарде возбуждение и сокращение в значительной степени перекрываются во времени, ПД заканчивается после начала фазы расслабления.
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 954; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.118.24.128 (0.143 с.) |