Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Понятие «регуляция внешнего дыхания»Стр 1 из 5Следующая ⇒
Понятие «регуляция внешнего дыхания»
Под регуляцией внешнего дыхания понимают: 1. генерация базового дыхательного паттерна (ритма) внешнего дыхания. 2. Изменение (модуляция) базового паттерна (ритма) внешнего дыхания в соответствии с метаболической потребностью организма вразных условиях[Мф2] жизнедеятельности.
Основные регулируемые параметры[Мф3]
Что регулируется? Объект регуляции?
1. Напряжение кислорода (pO2) в артериальной крови (95 —100 мм рт. ст.). 2. Напряжение углекислого газа (pCO2) в артериальной крови (в норме 38 —42 мм рт. ст.). 3. Регуляция МВЛ (МОД), т. е. глубины и частоты дыхания.
Основные виды (кибернетические) регуляции дыхания
1. по отклонению (отрицательная обратная связь с хеморецепторов при отклонении pO2, pCO2, pH артериальной крови); 2. по возмущению (например, влияние на дыхательный центр с проприорецепторов и моторной коры при физической работе); 3. по прогнозированию (например, условнорефлекторное влияние на дыхательный центр спортсмена-бегуна перед стартом).
Общая схема регуляции внешнего дыхания
Рис. 712280612. Общая схема регуляции внешнего дыхания.
Система регуляции дыхания (рис. 712280612) включает три основных элемента: 1) рецепторы, воспринимающие информацию и передающие ее в: 2) центральный регулятор, расположенный в головном мозге. Здесь информация обрабатывается и отсюда же посылаются команды на: 3) эффекторы (дыхательные мышцы), непосредственно осуществляющие вентиляцию легких.
Общую схему регуляции дыхания можно разбить на детали, например, представленные на рис. 712280632.
Рис. 712280632. Схему регуляции дыхания.
Центральный механизм дыхания
Включает: 1. дыхательные центры ствола головного мозга, 2. гипоталамус, 3. лимбическую систему, 4. кору больших полушарий.
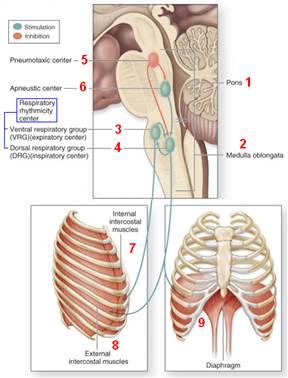
Дыхательные центры ствола головного мозга
Ствол мозга играет наиболее важную роль в регуляции дыхания. Автоматизм дыхания обусловлен зарождением импульсов в стволе головного мозга. Когда дыхание регулируется сознательно, кора головного мозга подчиняет себе эти центры автоматизма. Кроме того, при некоторых условиях в них могут поступать импульсы и от других отделов мозга. Но эти влияния ограничены.
В стволе выделяют дыхательные центры (рис. 712280709) 1. продолговатого мозга 1.1. вентролатеральный 1.2. дорсомедиальный 2. варолиева моста 2.1. пневмотаксический 2.2. апнейстический
Рис. 712280709. Дыхательные центры ствола головного мозга.
Чередование вдоха и выдоха обусловлено активностью нейронов, расположенных в варолиевом мосту и продолговатом мозге. Считается, что именно здесь находятся дыхательн ые цент ры. Они представляют собой не отдельные ядра, а довольно диффузные скопления нескольких групп нейронов.
Медуллярный дыхательный центр
Синонимы: бульбарный дыхательный центр.
Большой вклад в изучение его работы внесли М.Флуранс (1822 г.), Н.А. Миславский (1885 г.), Р.Баумгартен (1956 г.).
Расположен в ретикулярной формации в области дна IV желудочка у нижнего угла ромбовидной ямки
М едуллярный дыхательный центр состоит из групп нейронов локализованных в двух основных зонах продолговатого мозга: 1. в дорсомедиальных отделах (дорсальная дыхательная группа) и активизирующихся главным образом при вдохе. 2. в вентролатеральных отделах (вентральная дыхательная группа) и связана с выдохом и вдохом.
Раньше дорсальную дыхательную группу называли центром вдоха, а вентральную – выдоха. В настоящее время к таким обозначениям подходят осторожно.
Дыхательные нейроны
Дыхательные группы образованы дыхательными нейронами следующими основными типами (рис. 712280813):
Рис. 712280813
1. ранние инспираторные нейроны — интернейроны, которые активны в начале вдоха (расположены в вентролатеральнои группе); 2. полные инспираторные нейроны — эфферентные нейроны центра, иннервирующие мотонейроны мышц вдоха, активны в течение всего вдоха (расположены в дорсо-медиальной и вентролатеральнои группах); 3. поздние инспираторные нейроны — эфферентные нейроны центра, иннервирующие мотонейроны мышцвдоха и активные в конце вдоха (расположены в дорсомеди-
Некоторые авторы выделяют инспираторно‑тормозные нейроны — предполагаемые тормозные интернейроны инспираторной зоны, способные затормозить активность полных и поздних инспираторных нейронов.
4. постинспираторные нейроны — интернейроны, активные в первой половине выдохи, тормозящие как инспираторные, так и экспираторные нейроны (расположены в 5. экспираторные нейроны — эфферентные нейроны центра, иннервирующие мотонейроны мышц выдоха и активные во второй половине выдоха, преимущественно при 6. преинспираторные нейроны — интернейроны, блокирующие возбуждение экспираторных нейронов и способствующие смене выдоха на вдох.
Дорсальная дыхательная группа (ДДГ)
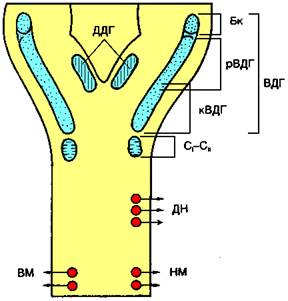
Рис. 712280818. Проекция местоположения дыхательного центра на дорсальную поверхность продолговатого мозга. ДДГ и ВДГ — соответственно дорсальная и вентральная дыхательные группы; Бк — комплекс Бётцингера; рВДГ и кВДГ — рост-ральняя м кяудальная часть ВДГ; СI—СII — сегменты спинного козгв; ДН, НМ и ВМ — соответственно димрртмальный нерв и нервы наружных и внутренних межреберных мышц.
Включает симметричные области продолговатого мозга, расположенные вентролатеральнее ядра одиночного пучка (рис. 712280818).
Дыхательные нейроны этой группы относятся только к инспираторному типу нейронов и представлены поздними и полными инспираторными нейронами.
Нейроны ДДГ получают афферентные сигналы от легочных рецепторов растяжения по волокнам блуждающего нерва, нейроны которого имеют обширные синаптические связи с другими отделами дыхательного центра и с различными отделами ЦНС. Только часть инспираторных нейронов ДДГ связана аксонами с дыхательными мотонейронами спинного мозга, преимущественно с контралатеральной стороны.
Вентральная дыхательная группа (ВДГ)
Вентральная дыхательная группа (ВДГ) расположена латеральнее обоюдного ядра продолговатого мозга, или ядра блуждающего нерва. ВДГ подразделяется на ростральную и кау-дальную части относительно уровня задвижки (obex) продолговатого мозга (см. рис. 712280818). Ростральная часть ВДГ состоит из инспираторных нейронов разных типов: ранних, полных, поздних инспираторных и постинспи-раторных. Ранние инспираторные и постинспираторные нейроны ВДГ называются проприобульбарными нейронами, так как они не направляют свои аксоны за пределы дыхательного центра продолговатого мозга и контактируют только с другими типами дыхательных нейронов. Часть полных и поздних инспираторных нейронов направляют свои аксоны к дыхательным мотонейронам спинного мозга, а следовательно, управляют мышцами вдоха. Каудальная часть ВДГ состоит только из экспираторных нейронов. Все экспираторные нейроны направляют аксоны в спинной мозг. При этом 40% экспираторных нейронов иннервирует внутренние межреберные мышцы, а 60% — мышцы брюшной стенки. Ростральнее ВДГ локализованы компактной группой экспираторные нейроны (комплекс Бетцингера), аксоны которых связаны только с другими типами нейронов дыхательного центра. Предполагают, что именно эти нейроны синхронизируют деятельность правой и левой половин дыхательного центра.
В непосредственной близости от нейронов ВДГ расположены различные типы респираторно-связанных нейронов, которые иннер-вируют мышцы верхних дыхательных путей и гортани. Нейроны дыхательного центра в зависимости от проекции их аксонов подразделяют на три группы: 1) нейроны, иннервирующие мышцы верхних дыхательных путей и регулирующие поток воздуха в дыхательных путях; 2) нейроны, которые синаптически связаны с дыхательными мотонейронами спинного мозга и управляют таким образом мышцами вдоха и выдоха; 3) проприобульбарные нейроны, которые связаны с другими нейронами дыхательного центра и участвуют только в генерации дыхательного ритма.
Довольно распространено (хотя и не общепринято) мнение о том, что нейроны инспираторной зоны способны к самопроизвольному периодическому возбуждению, и именно они отвечают за периодичность дыхания. При устранении всех возможных афферентных стимулов эти инспираторные нейроны продолжают в определенном ритме генерировать залпы потенциалов действия, которые передаются к диафрагме и другим инспираторным мышцам. Очередное возбуждение нейронов инспираторной зоны начинается после латентного периода (т.е. периода отсутствия активности) длительностью в несколько секунд. Затем появляются потенциалы действия, и частота их в следующие секунды экспоненциально увеличивается. Соответственно этому нарастает и активность инспираторных мышц. Затем генерация потенциалов действия в инспираторной зоне прекращается, и тонус этих мышц снижается до исходного уровня.
Инспираторная фаза Соответствует вдоху. Обусловлена последовательной активацией ранних, полных и поздних инспираторных нейронов, что сопровождается линейным нарастанием их суммарной активности. Резкое уменьшение их активности (смена вдоха на выдох), как полагают, связано с
Постинспираторная фаза Соответствует первой половине выдоха (пассивная экспирация) и обусловлена особыми постинспираторными нейронами, которые тормозят как инспираторные, так и экспираторные нейроны. Они, вероятно, обеспечивают интервал времени, необходимый для выведения воздуха из легких за счет их эластической тяги;
Экспираторная фаза Соответствует второй половине В конце выдоха происходит возбуждение преинспираторных нейронов, которые тормозят импульсацию экспираторных нейронов (прекращают выдох). Возможным источником возбуждения преинспираторных нейронов являются ирритантные рецепторы легких, возбуждающиеся при уменьшении объема легких во время выдоха (инспираторно-облегчающий рефлекс Геринга—Брейера). При частом дыхании экспираторная фаза может быть не выражена, и постинспираторная фаза непосредственно переходит в следующую фазу инспирации.
Гипоталамус Осуществляет связь дыхания с обменом веществ и терморегуляцией в организме. Регулирует дыхание для обеспечения поведенческих актов, направленных на удовлетворение биологических потребностей (агрессивно-оборонительной, пищевой, половой и др.).
Лимбическая система.
Осуществляет связь дыхания с вегетативной регуляцией внутренних органов и эмоциями.
Кора больших полушарий. По пирамидным путям, минуя дыхательный центр, оказывает влияние непосредственно на спинальные моторные центры дыхательных мышц (поэтому при некоторых поражениях
Осуществляет условнорефлекторную и произвольную регуляцию дыхания. Осуществляет корковое дублирование автоматии дыхательного центра (например, при поражении периодической деятельности дыхательного центра — синдроме Ундины). Регулирует дыхание для обеспечения социальных форм поведения.
Синдром "Проклятие Ундины" ("La Malediction d'Ondine") встречающийся при нарушении работы дыхательного центра. При нем человек вообще не может самостоятельно дышать во время сна. Пациента (днем - вполне здорового человека) подключают на ночь к аппарату искусственной вентиляции легких! Считается самым тяжелым видом апноэ (отсутствие дыхания). В основу названия положена легенда, согласно которой водяная фея Ундина, обманутая мужем, лишает его всех автоматических функций, по некоторым данным с помощью затяжного поцелуя. С этого момента он должен постоянно помнить, что ему нужно дышать, держать в поле внимания акт ходьбы, все действия руками и т. д. Заснув, он умирает, так как перестает управлять волевыми усилиями дыхательным центром и другими жизненно важными функциями
Рецепторы растяжения легких Медленно адаптируются. Расположены в гладких мышцах трахеи и бронхах, реагируют на увеличение объема легких при вдохе; с них возникает инспираторно-тормозящий рефлекс Геринга—Брейера (если дыхательный объем более 1 л);
Ирритантные рецепторы Быстро адаптирующиеся рецепторы, порог раздражения которых выше, чем рецепторов растяжения легких;
Расположены в эпителии бронхов, реагируют на быстрое изменение объема легких, на механические воздействия (пыль) и пары химических веществ: С
Рефлексы Геринга — Брейера.
Раздувание легких у наркотизированного животного рефлекторно тормозит вдох и вызывает выдох. Нервные окончания, расположенные в бронхиальных мышцах, играют роль рецепторов растяжения легких. Их относят к медленно адаптирующимся рецепторам растяжения легких, которые иннервируются ми-елинизированными волокнами блуждающего нерва. Рефлекс Геринга — Брейера контролирует глубину и частоту дыхания. У человека он имеет физиологическое значение при дыхательных объемах свыше 1 л (например, при физической нагрузке). У бодрствующего взрослого человека кратковременная двусторонняя блокада блуждающих нервов с помощью местной анестезии не влияет ни на глубину, ни на частоту дыхания. У новорожденных рефлекс Геринга — Брейера четко проявляется только в первые 3—4 дня после рождения.
Проприоцептивный контроль дыхания. Рецепторы суставов грудной клетки посылают импульсы в кору больших полушарий и являются единственным источником информации о движениях грудной клетки и дыхательных объемах. Межреберные мышцы, в меньшей степени диафрагма, содержат большое количество мышечных веретен. Активность этих рецепторов проявляется при пассивном растяжении мышц, изометрическом сокращении и изолированном сокращении интрафузальных мышечных волокон. Рецепторы посылают сигналы в соответствующие сегменты спинного мозга. Недостаточное укорочение инспираторных или экспираторных мышц усиливает импульсацию от мышечных веретен, которые через у-мотонейроны повышают активность о-мотонейронов и дозируют таким образом мышечное усилие. Хеморефлексы дыхания. Рог и Рсог в артериальной крови человека и животных поддерживается на достаточно стабильном уровне, несмотря на значительные изменения потребления Оз и выделение С02. Гипоксия и понижение рН крови (ацидоз) вызывают усиление вентиляции (гипервентиляция), а гипероксия и повышение рН крови (алкалоз) — понижение вентиляции (гиповентиляция) или апноэ. Контроль за нормальным содержанием во внутренней среде организма 02, СОг и рН осуществляется периферическими и центральными хеморецепторами. Адекватным раздражителем для периферических хеморецепторов является уменьшение Ро; артериальной крови, в меньшей степени увеличение Рсо2 и рН, а для центральных хеморецепторов — увеличение концентрации Н* во внеклеточной жидкости мозга. Артериальные (периферические) хеморецепто-ры. Периферические хеморецепторы находятся в каротидных и
аортальных тельцах. Сигналы от артериальных хеморецепторов по синокаротидным и аортальным нервам первоначально поступают к нейронам ядра одиночного пучка продолговатого мозга, а затем переключаются на нейроны дыхательного центра. Ответ периферических хеморецепторов на понижение Рао^ является очень быстрым, но нелинейным. При Рао; в пределах 80—60 мм рт. ст. (10,6—8,0 кПа) наблюдается слабое усиление вентиляции, а при Рао; ниже 50 мм рт. ст. (6,7 кПа) возникает выраженная гипервентиляция. Расо2 и рН крови только потенцируют эффект гипоксии на артериальные хеморецепторы и не являются адекватными раздражителями для этого типа хеморецепторов дыхания. Реакция артериальных хеморецепторов и дыхания на гипоксию. Недостаток С>2 в артериальной крови является основным раздражителем периферических хеморецепторов. Импульсная активность в афферентных волокнах синокаротидного нерва прекращается при Раод выше 400 мм рт. ст. (53,2 кПа). При нормоксии частота разрядов синокаротидного нерва составляет 10% от их максимальной реакции, которая наблюдается при Раод около 50 мм рт. ст. и ниже-Гипоксическая реакция дыхания практически отсутствует у коренных жителей высокогорья и исчезает примерно через 5 лет у жителей равнин после начала их апаптации к высокогорью (3500 м и выше). Центральные хеморецепторы. Окончательно не установлено местоположение центральных хеморецепторов. Исследователи считают, что такие хеморецепторы находятся в ростральных отделах продолговатого мозга вблизи его вентральной поверхности, а также в различных зонах дорсального дыхательного ядра. Наличие центральных хеморецепторов доказывается достаточно просто: после перерезки синокаротидных и аортальных нервов у подопытных животных исчезает чувствительность дыхательного центра к гипоксии, но полностью сохраняется реакция дыхания на гиперкапнию и ацидоз. Перерезка ствола мозга непосредственно выше продолговатого мозга не влияет на характер этой реакции. Адекватным раздражителем для центральных хеморецепторов является изменение концентрации Н4 во внеклеточной жидкости мозга. Функцию регулятора пороговых сдвигов рН в области центральных хеморецепторов выполняют структуры гематоэнцефали-ческого барьера, который отделяет кровь от внеклеточной жидкости мозга. Через этот барьер осуществляется транспорт 02, С02 и Н^ между кровью и внеклеточной жидкостью мозга. Транспорт СОз и H+ из внутренней среды мозга в плазму крови через структуры гематоэнцефалического барьера регулируется с участием фермента ка рбоангидразы. Реакция дыхания на COi. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз тормозят центральные хеморецепторы. Для определения чувствительности центральных хеморецепторов к изменению рН внеклеточной жидкости мозга используют метод возвратного дыхания. Испытуемый дышит из замкнутой емкости, заполненной предварительно чистым Од. При дыхании в замкнутой
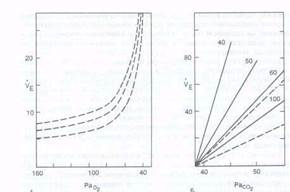
Рис. 8.12. Изменение вентиляции легких (ve. л'мин) в зависимости от парциального давления Од (А) и COz <Б) в альвеолярном воздухе при различном содержании Ог в альвеолярном воздухе (40, 50. 60 и 100 мм рт. ст.). системе выдыхаемый СО; вызывает линейное увеличение концентрации СОа и одновременно повышает концентрацию Н* в крови, а также во внеклеточной жидкости мозга. Тест проводят в течение 4—5 мин под контролем содержания СО; в выдыхаемом воздухе- На рис. 8.12 показано изменение объема вентиляции при различном уровне напряжения С02 в артериальной крови. При Расоа ниже 40 мм рт. ст. (5,3 кПа) может возникнуть апноэ в результате гипокапнии. В этот период дыхательный центр мало чувствителен к гипоксической стимуляции периферических хеморецепторов. 8.6.3. Координация дыхания с другими функциями организма В филогенетическом развитии организма человека и животных дыхательный центр приобретает сложные синаптические взаимоотношения с различными отделами ЦНС. В отличие от других физиологических функций организма дыхание находится под контролем автономной (вегетативной) и соматической нервной системы, поэтому у человека и животных дыхание нередко называют вегето-сомати ческой функцией. Существует тесное взаимодействие регуляции дыхания гуморальной и рефлекторной природы и процессами сознательной деятельности мозга. Однако во время сна или в состояниях, связанных с отсутствием сознания у человека, сохраняется внешнее дыхание и обеспечивается нормальное поддержание газового гомеостаза внутренней среды. С другой стороны, человек имеет возможность по собственному желанию
изменять глубину и частоту дыхания или задерживать его, например во время пребывания под водой. Произвольное управление дыханием основано на корковом представительстве проприоцептивного анализатора дыхательных мышц и на наличии коркового контроля дыхательных мышц. Электрическое раздражение коры больших полушарий у человека и животных показало, что возбуждение одних корковых зон вызывает увеличение, а раздражение других — уменьшение легочной вентиляции. Наиболее сильное угнетение дыхания возникает при электрической стимуляции лимбической системы переднего мозга. При участии центров терморегуляции гипоталамуса возникает ги-перпноэ при гипертермических состояниях. Однако многие нейрофизиологические механизмы взаимодействия нейронов переднего мозга с дыхательным центром остаются пока мало изученными. Дыхание опосредованно через газы крови влияет на кровообращение во многих органах. Важнейшим гуморальным, или метаболическим, регулятором локального мозгового кровотока являются Н* артериальной крови и межклеточной жидкости. В качестве метаболического регулятора тонуса сосудов мозга рассматривают также С02. В последнее время эта точка зрения подвергается сомнению, поскольку СО-1 как молекулярное соединение практически отсутствует во внутренней среде организма. Молекулярный С02 (0—С=-0) встречается в организме в альвеолярном воздухе, а в тканях только при переносе СОд через аэрогематический и гистогематический барьеры. В крови и межклеточной жидкости СО; находится в связанном состоянии, в виде гидрокарбонатов, поэтому правильнее говорить о метаболической регуляции Н^ тонуса гладких мышц артериальных сосудов и их просвета. В головном мозге повышение концентрации Н^ расширяет сосуды, а понижение концентрации I^ в артериальной крови или межклеточной жидкости, напротив, повышает тонус гладких мышц сосудистой стенки. Возникающие при этом изменения мозгового кровотока способствуют изменению градиента рН по обе стороны гематоэнцефалического барьера и создают благоприятные условия либо для вымывания из сосудов мозга крови с низким значением рН, либо для понижения рН крови в результате замедления кровотока. Функциональное взаимодействие систем регуляции дыхания и кровообращения является предметом интенсивных физиологических исследований. Обе системы имеют общие рефлексогенные зоны в сосудах: аортальную и синокаротидные. Периферические хеморецепторы дыхания аортальных и каротидных телец, чувствительные к гипоксии в артериальной крови, и барорецепторы стенки аорты и каротидных синусов, чувствительные к изменению системного артериального давления, расположены в рефлексогенных зонах в непосредственной близости друг от друга. Все названные рецепторы посылают афферентные сигналы к специализированным нейронам основного чувствительного ядра продолговатого мозга — ядра одиночного пучка. В непосредственной близости от этого ядра находится дорсальное дыхательное ядро дыхательного центра. Здесь же в продолговатом мозге находится сосудодвигательный центр. Координацию деятельности дыхательного и сосудодвигательного центров продолговатого мозга осуществляют нейроны ряда интег-ративных ядер бульбарной ретикулярной формации.
Понятие «регуляция внешнего дыхания»
Под регуляцией внешнего дыхания понимают: 1. генерация базового дыхательного паттерна (ритма) внешнего дыхания. 2. Изменение (модуляция) базового паттерна (ритма) внешнего дыхания в соответствии с метаболической потребностью организма вразных условиях[Мф2] жизнедеятельности.
Основные регулируемые параметры[Мф3]
Что регулируется? Объект регуляции?
1. Напряжение кислорода (pO2) в артериальной крови (95 —100 мм рт. ст.). 2. Напряжение углекислого газа (pCO2) в артериальной крови (в норме 38 —42 мм рт. ст.). 3. Регуляция МВЛ (МОД), т. е. глубины и частоты дыхания.
|
|||||||||||
|
|
Последнее изменение этой страницы: 2016-09-17; просмотров: 520; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.221.111.22 (0.102 с.) |




 них формируются рефлексы кашля, сужения бронхов при выдохе, инспираторно-облегчающий рефлекс Геринга — Брейера при спадении легких, который укорачивает выдох и способствует его смене на вдох;
них формируются рефлексы кашля, сужения бронхов при выдохе, инспираторно-облегчающий рефлекс Геринга — Брейера при спадении легких, который укорачивает выдох и способствует его смене на вдох;