Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Системный подход в гидроэкологических исследованиях
Как уже упоминалось ранее, водные экосистемы – основной объект изучения в гидроэкологии. Экосистемное направление, пожалуй, лучше всего демонстрирует принципы системности в экологической науке. Экологическая система представляет собой любую совокупность живых оpганизмов и сpеды их обитания, взаимосвязанных обменом веществ, энеpгии, и инфоpмации, котоpую можно огpаничить в пpостpанстве и во вpемени по значимым для конкpетного исследования пpинципам. Любая экологическая система является открытой, поскольку она всегда взаимодействует с внешней средой: солнечной радиацией, влагообоpотом на поверхности и в грунтах, ветровым переносом и выносом материала. Следовательно, любые пространственные ограничения экосистемы всегда условны. Понятие экологической системы иерархично. Это означает, что всякая экологическая система определенного уровня включает в себя ряд экосистем предыдущего уровня, меньших по площади и сама она, в свою очередь, является составной частью более крупной экосистемы. Например, правомерно рассматривать в качестве экосистемы озерную впадину, ограниченную склонами котловины. Продолжая этот ряд вверх, можно подойти к экологической системе Земли – биосфере, а двигаясь вниз – к биогеоценозу, как элементарной биохорологической единице биосферы. Любая экосистема состоит из биотических (живые организмы) и абиотических (косная или неживая природа) компонентов. Экологические компоненты – основные материально-энергетические составляющие экосистемы (В.В. Дмитриев курс лекций «Современные проблемы экологии и природопользования», 2011). В совокупности компоненты экосистемы определяют и лимитируют ее развитие. К биотическим компонентам экосистемы, в том числе и водной, относятся продуценты, консументы и редуценты, которые связаны друг с другом через биогенные компоненты экосистемы – живое и мертвое органическое вещество. Абиотические компоненты весьма разнообразны, к ним относятся и морфологические характеристики водного объекта, и метеорологические условия, динамические и химические факторы, и ряд других. Все эти факторы тесно связаны друг с другом и в комплексе создают все разнообразие мест обитания, которое и отражается в распределении гидробионтов в водоемах (В.В. Дмитриев курс лекций «Современные проблемы экологии и природопользования, 2011, с. 268).
Абиотическая составляющая водных экосистем является объектом изучения различных направлений гидрометеорологического цикла наук. Гидрофизика, гидромеханика, гидродинамика, гидрохимия, метеорология – все эти науки изучают процессы, составляющие абиотические факторы водной среды. Гидроэкология (а также гидробиология) изучает взаимодействие этих факторов с биотической составляющей водной экосистемы, используя при этом законы и методы других наук. При этом биотические факторы «накладываются» на абиотическую составляющую. Более наглядно эти идеи будут показаны в главе про моделирование. Моделирование как метод исследований в гидроэкологии и науке вообще хотя и возник задолго до распространения идей системного подхода, во многом опирается на него и является наглядной демонстрацией сущности системного подхода.
Системный подход к изучению любого объекта, например экосистемы, состоит, во-первых, в определении образующих ее составных частей — Х 1,..., Хnи взаимодействующих с ней объектов окружающей среды — S 1,…, Sk, во-вторых, в установлении структуры экосистемы, т. е. совокупности внутренних связей и отношений σ1,…,σ r, а также связей между экосистемой и окружающей средой и, в-третьих, в нахождении функции (закона функционирования) экосистемы F, определяющей характер изменения компонентов экосистемы и связей между ними под действием внешних объектов S 1(t),…, Sk (t). Для решения этих трех основных задач в арсенале современной экологии имеются разнообразные методы исследования, среди которых выделяются три основные группы методов: 1) полевые наблюдения; 2) эксперименты в поле и в лаборатории; 3) моделирование. Каждый из них имеет свою историю развития, отражающую последовательные этапы углубления знаний об экосистемах, и соответствует вполне определенной стадии единого процесса системного экологического исследования. Остановимся на каждом из этих методов в отдельности, а затем рассмотрим, как они интегрируются в рамках системного подхода к изучению экосистем.
Полевые наблюдения Метод непосредственного наблюдения изучаемой экосистемы пли ее определенных компонентов в естественных условиях подразумевает невмешательство (или минимально возможное вмешательство) наблюдателя и представляет собой наиболее важный, исторически первый прием экологического исследования (в так называемом «интактном» режиме). Известный так же как сравнительный эколого-географический метод (Кашкаров, 1938), он прошел длительный путь развития от красочных, порой романтических, описании картин природы, вышедших из-под пера выдающихся натуралистов своего времени, таких как Л. Гумбольдт, Ч. Дарвин, Л. Уоллес, Н. Л. Северцов и др., до современных комплексных программ экологических наблюдений и измерений с помощью новейшей электронной, акустической, фотографической и прочен аппаратуры, при осуществлении стационарных исследований экосистем. Полевые исследования экосистем некоторого ландшафта призваны разрешить следующие задачи. 1. Выделение основных типов экосистем и их взаимосвязей в изучаемом ландшафте. 2. Определение видового состава организмов, населяющих каждую из экосистем, установление соответствующего ей микроклимата, типа почвы, почвообразующей породы, характера гидрологического режима., 3. Идентификация структуры экосистемы на качественном уровне, т. е. получение общей картины отношений между видами, установление характера связен организмов с почвой, приземным слоем воздуха и другими неживыми компонентами экосистемы, а также этих последних друг с другом. 4. Получение количественных оценок для основных показателей состава экосистемы, например определение плотностей популяций, нахождение (установление) значений основных переменных состояния микроклимата (температуры, влажности, концентрации СО2 и др.), почвы (температуры, влажности, концентрации элементов питания растений п т. п.), для наземных экосистем и водной массы (температуры, солености, концентрации 02, ионов водорода, биогенных элементов и т. д.), для водных экосистем. 5. Количественная идентификация структуры экосистемы, т. е. количественное описание функциональных связей между компонентами системы и внешних воздействий на систему[1]. Примером здесь может служить установление зависимости: 1) интенсивности Фотосинтеза от освещенности, температуры, влажности, обеспеченности биогенными элементами и т. д.; 2) выяснение зависимости скорости выедания растений растительноядными животными от наличного запаса и качества фитомассы, от плотности и состояния самой популяции растительноядных, от метеорологических условий и других факторов; 3) скорости испарения воды из почвы от метеоусловий, свойств почвы, растительности и т. д. 6. Комплексное описание сопряженной динамики всех компонентов экосистемы в сезонном, годовом и многолетнем плане, которое могло бы служить основой для глубокого анализа закономерностей функционирования данной экосистемы и ее сравнения с другими экосистемами. Несмотря на отсутствие в современной экологии общепринятой классификации экосистем для ландшафтов всех природных зон уже известны наиболее распространенные типы слагающих их экосистем и сформулированы основные принципы их выделения. Руководствуясь ими, эколог может уже в поле выделить экосистемы и наметить их границы, а при последующей камеральной обработке, опираясь на дополнительные данные о геоморфологии, составить развернутое описание и дать полное наименование всех выделенных экосистем.
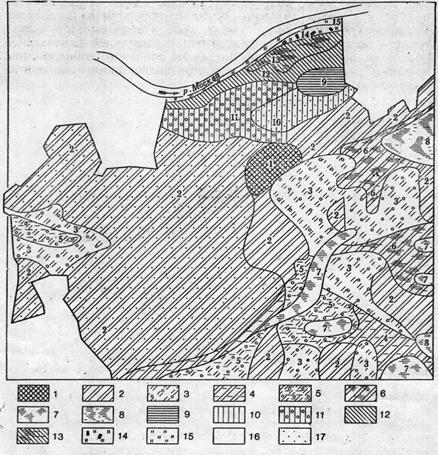
Рис. IV.2. Карта растительности территории Звенигородской биостанции МГУ (Барсукова, Пятковская, 1962) Еловые леса: 1 —ельники; 2— елово-сосновые леса; 3 — елово-сосново-березовыс леса; 4 — елово-березовые леса; 5 — сложные ельники; 6 — елово-березозый лес с подлеском. Сосновые леса: 7 — сосняки; 8 — сосново-еловые леса; 9 — сосново-елово-березовые леса; 10 — сосноео-березовые леса. Мелколиственные леса: 11 — березняки; 12 — беролово-елозыо леса; 13 — березово-сосновке леса; 14 — берсзово-сосиотю-еловые леса; 15 — берсзово-сосново-елово-осиновые леса; 16 — березово-сосново-осиновые леса; 17 — березово-осиновые леса; 18— бере-зоао-липовые леса; 19— осинники; 20 — ольшаники; 21 — смешанные леса; 22 — луга; 23—-лесные поляны; 24 — болото
Трудности, характерные для этого этапа полевых исследований, можно показать на следующем примере. На рис. IV.2 представлена подробная карта растительности Звенигородской биостанции Московского университета, территория которой охватывает такие типичные для ландшафтов средней полосы геоморфологические элементы, как ровные водораздельные пространства, склоны, террасы, пойма реки и сама река (рис. IV.З). В общей сложности на карте показано 24 типа различных экосистем, перечисленных в легенде (рис. IV.2). Высокая пестрота растительного покрова, многочисленность экосистем, находящихся на разных стадиях вторичной сукцессии, искусственность границ, во многих случаях совпадающих с прямоугольными линиями лесоустроительных участков или дорогами, отражают сложную историю длительного воздействия человека на экосистемы рассматриваемого ландшафта. Однако, обратившись к почвенной карте той же территории (рис. IV.4), мы обнаруживаем заметно менее пеструю картину (количество выделяемых учетных единиц падает с 24 до 15, еще сильнее снижается число конкретных выделов), на которой
Рис. IV.З. Ландшафтно-геоморфологический профиль северного участка территории Звенигородской биостанции МГУ от водораздела к р. Москве (по Барсуковой, Пятковcкой, 1962)
Рис. IV.4. Почвенная карта Звенигородской биостанции биологического факультета МГУ (составлена К. М. Смирновой, 1962)
Почвы водораздела. 1 — подзол; 2 — сильноподзолистые; 3 — дерново-сильноподзолистые; 4 — сильноподзолистые слабоглинистые; 5 — дерново-сильноподзолистые слабоглинистые; 6 — сильноподзолистые слабоглинистые в комплексе с торфянисто-подзолистыми сильноглинистыми; 7 — торфянисто-перегнойно-глеевые; 8— торфяно-перегнойно-глеевые. Почвы террас р. Москвы. III терраса: 9 — дерново-среднеподзолистые; • 10 — бурые лесные слабооподзоленные; 11 — бурые лесные сильнооподзоленные; II терраса: 12 — дерново-сильноподзолистые; I терраса: 13 — дерново-сильноподзолистые в комплексе с перегнойно-глеевыми; пойменная терраса: 14 — дерново-луговые слабокарбонатные на погребенных дерново-подзолистых; 15 — дерново-луговые карбонатные. Механический состав почв: 16 — суглинистый; 17 — песчаный хорошо проявляется закономерное размещение почв и соответствовавших им в прошлом коренных экосистем, обусловленных прежде всего геоморфологическим строением территории (водораздельное пространство, терраса и пойма р. Москвы) и механическим составом поверхностных отложений (песчаных или суглинистых). Кроме необходимости учета истории антропогенного воздействия при идентификации состава экосистем изучаемого ландшафта приходится решать много других вопросов, связанных с явлениями непрерывности (континуальности) пространственного изменения абиотических факторов и биоты некоторых ландшафтов, с процессами миграции популяций, сукцессионными изменениями и др. Поэтому первоначальное экологическое подразделение ландшафта на элементарные экосистемы редко бывает окончательным. Как правило, оно уточняется в ходе последующих исследований. После выделения основных типов экосистем и установления их взаимосвязей в ландшафтной системе внимание исследователей сосредоточивается на изучении состава, структуры и функционирования каждой экосистемы с тем, чтобы на основе этих данных подойти к более глубокому пониманию функционирования изучаемого ландшафта в целом. Изложению методики полевых экологических исследований посвящены труды таких известных ученых, как А. Тэнсли (Tanslеу, 1923, 1946); Ч. Элтон (Elton, 1927; Elton, Miller, 1954), В. Шелфорд (Shelford, 1929), В. Н. Сукачев (1930), Л. Г. Раменский (1937, 1938), Ф. Клементс и В. Шелфорд (Clements, Shelford, 1939), Г. А. Новиков (1949) и др. Однако при всех достоинствах названных руководств они адресованы прежде всего индивидуальным исследователям или небольшим коллективам, ориентированным на специальное изучение той или иной группы организмов. Обладая весьма скромными средствами, такие коллективы могли получить лишь весьма фрагментарные сведения о многообразии и динамичности экологических процессов, которые было трудно использовать для построения целостной картины всей экосистемы. Хотя многие из перечисленных авторов сознательно пропагандировали методологию, которую мы сегодня называем «системным подходом», до конца 50-х гг. XIX в. в среде экологов все еще господствовала установка на узкоспециализированные исследования, когда, как это образно заметил Л. Оман (Hauman, 1933), каждый ученый был занят «терпеливой и привычной эксплуатацией своей индивидуальной» жилы, несмотря на то что «пустая порода уже заполнила полость шахты». Попытки объединения под флагом так называемых «комплексных» исследований разных специалистов, которые, однако, не были связаны общей теоретической и методологической концепцией (каковыми позднее явились концепция экосистемы или биогеоценоза), неоднократно предпринимавшиеся как в нашей стране, так и за рубежом, приносили, как правило, весьма скудные (с точки зрения системного подхода) результаты. В лучшем случае они завершались публикацией очередного сборника, в котором мирное и независимое сосуществование статей по ботанике, зоологии, микробиологии, почвоведению, метеорологии и другим дисциплинам лишь изредка нарушалось робкими попытками междисциплинарного синтеза с целью получения действительно комплексного описания экосистемы во всей ее целостности.
Существенный прогресс в развитии экологических исследований на основе системного подхода, включая долгосрочные комплексные наблюдения, начинается в конце 50-х — начале 60-х гг. Он связан с работами Говарда Одума (Odum H., 1956, 1957, 1960), В. Н. Сукачева и его сотрудников (Сукачев и др., 1957; Сукачев, Зонн, 1961) и ряда других зарубежных исследователей. Всесторонние экологические исследования приобрели -широкий размах на стационарах Международной биологической программы (Программа и методика..., 1966, 1974). Однако наивысшим достижением в области полевых экологических наблюдений являются программы комплексного экологического мониторинга, развертываемые на биосферных заповедниках в соответствии с проектом организации Глобальной системы мониторинга окружающей среды (ГСМОС). Эти программы предусматривают проведение всесторонних наблюдений над -динамикой наиболее характерных типов целинных экосистем во всех основных биотических провинциях, результаты которых в сочетании с материалами интенсивных исследований на специальных полигонах и опытных станциях предполагается использовать для построения математических моделей этих экосистем, обеспечивающих возможность прогнозирования и оптимального управления их функционированием (UNESCO, 1972; UNESCO and UNEP, 1974; «Биосферные заповедники», 1977).
Эксперимент Наряду с наблюдением широкое применение в экологии, как и в других естественных науках, имеет экспериментальный метод. Принципиальное отличие эксперимента от пассивного наблюдения заключается в том, что задача наблюдателя — получение сведений о поведении экосистемы в ненарушенных естественных условиях (точнее, при минимально возможных возмущениях, вызываемых процессом наблюдения), тогда как в эксперименте исследователь следит за экосистемой, в которой им сознательно произведены определенные (быть может, и достаточно сильные) изменения. Поэтому, когда эколог с помощью сложной аппаратуры следит за перемещением стада оленей, некоторые из которых снабжены миниатюрными радиопередатчиками, то при этом он занимается наблюдениями, а не экспериментами. Напротив, регистрация динамики численности того же стада пусть даже самыми приближенными методами, без всякой аппаратуры, но после предварительной элиминации из экосистемы хищников, например волков, либо после введения искусственной подкормки в суровых зимних условиях, будет настоящим экологическим экспериментом. Многообразие экспериментов в экологии столь же необозримо, как необозримо число возможных воздействий экспериментатора на экосистему. Поэтому весьма полезным оказывается выделение различных категорий экологических экспериментов в зависимости от степени контроля человека над изучаемой экосистемой и числа сознательно варьируемых факторов. Прежде всего, целесообразно различать эксперименты" по достигнутому в них уровню контроля над объектом изучения. На одном конце спектра располагаются практически неконтролируемые эксперименты, когда, произведя в начале опыта некоторое воздействие на экосистему, экспериментатор впоследствии только следит за ее динамикой, которая развертывается на фоне всевозможных, часто непредвиденных и нежелательных, с точки зрения экспериментатора, внешних воздействий, например погодных. На противоположном конце спектра группируются эксперименты, в которых исследователь имеет полную возможность контролировать все интересующие его экологические факторы в течение всего времени опыта. Промежуточное положение занимают эксперименты, в которых могут контролироваться лишь некоторые факторы. Вспоминая широко распространенное деление экспериментов на полевые и лабораторные, можно заметить, что полевые эксперименты в большинстве случаев относятся к неконтролируемым, так как возможности исследователя контролировать экологические факторы в природных условиях очень и очень ограничены. В лабораторных экспериментах можно обеспечить контроль большего числа факторов и, тем не менее, большинство лабораторных опытов также относится к категории частично контролируемых. Сколько-нибудь полный охват всех существенных факторов достигается только на сложных и дорогостоящих экспериментальных установках, наподобие фитотронов, акватронов и других управляемых лабораторных «микрокосмов». Классической схемой проведения естественнонаучных экспериментов со времени Ф. Бэкона считается так называемый однофакторный эксперимент, когда изучается влияние одного избранного фактора при фиксированных значениях всех остальных. Но если в науках о неживой природе (и прежде всего в физике) однофакторные эксперименты оказались весьма плодотворными и принесли немало выдающихся открытий, то в экспериментах с биологическими объектами, поведение которых существенно зависит от целого комплекса факторов, как, например, природные или сельскохозяйственные экосистемы, этот подход оказался менее эффективным. Для решения многофакторных задач, связанных с получением высоких урожаев, в 30-х гг. нашего столетия Ренальдом Фишером были разработаны основы метода многофакторных экспериментов, состоящего в том, что в каждом варианте из серий опытов экспериментатор изменяет не один, а сразу несколько факторов, значения которых комбинируются определенным способом, в результате чего при последующей математической обработке оказывается возможным получить искомое многофакторное описание изучаемого процесса или явления. Эффективность применения метода планирования многофакторных экспериментов в экологических исследованиях убедительно показана в работах отечественных авторов (Федоров, 1967; Максимов, Федоров, 1967, 1969), которые предложили использовать планы факторного эксперимента при исследовании связи первичной продукции с уровнями факторов, действующих в водоемах. Предложенный метод планируемых добавок, представляющий собой комбинацию обычного метода подкормок Шрайбера, например биогенных элементов на активность фитопланктона, и планов факторного эксперимента, позволил получать информацию (качественную и количественную) о характере взаимодействий факторов системы. В этом случае факторы, включенные в план эксперимента, меняются одновременно и независимо друг от друга, по определенным планам, и по результатам их влияния на величину первичной продукции рассчитываются соответствующие значения bi как при линейных членах (bi), так и при их взаимодействиях (bij). В уравнении модели вида

где xi,- — переменные (факторы), а bi,- — коэффициент регрессии при соответствующих переменных, значения которых определяют величину отклика в изучаемой области. Таким образом, уравнение (IV.6) связывает величину первичной продукции (y) с уровнями (концентрациями) факторов, действующих в системе. Проведенные В. Д. Федоровым и его сотрудниками исследования у Карельского побережья Белого моря позволили исследовать зависимость первичной продукции от освещенности, концентрации ряда биогенных элементов и состава фитопланктона в водоеме. Полученные количественные оценки при обнаруженных взаимодействиях факторов позволили достаточно подробно исследовать связь первичной продукции и концентрации в мирской воде нитратного и аммиачного азота, фосфора, железа и кремния (Федоров, Белая, Максимов, 1970; Федоров, Семин, 1970). Заметную роль в развитии экологии сыграли эксперименты по лабораторному культивированию отдельных популяций или целых микроэкосистем. Хотя первые исследования такого рода проводились еще в XIX в., как, например, известные работы Р. Уорингтона но сбалансированным аквариумам (Warington, 1851, 1857), широкое использование лабораторных микроэкосистем для решения сннэкологических (в отличие от экофизиологических и популяционно-биологических) задач началось лишь в 30-х гг. нашего века. Успешному развертыванию этих исследований, без сомнения способствовал опыт, накопленный в серии блестящих работ по лабораторному культивированию отдельных популяций, выполненных в 20-х гг. Р. Пирлом с сотрудниками (Pearl, Parker, 1922; Pearl, 1925, 1928), В. Олли (Allee, 1926, 1927), Р. Чепмэном (Chapman, 1928), И. И. Калабуховым (1929). Но если натканные исследователи в своих экспериментах работали с отдельными популяциями, определяли такие характеристики, как рождаемость, смертность, продолжительность жизни, форма кривой роста и т. п., в скнэкологическнх и «микрокосменных» экспериментах изучалось взаимодействие нескольких популяций и целостные свойства экосистемы. В области лабораторных синэкологических экспериментов выдающееся значение имеют исследования, проводившиеся в начале 30-х гг. в нашей стране под руководством В. В. Алпатова (Алпатов, 1931, 1932, 1934; Алпатов и Гаузе, 1933)} и в особенности серия классических работ Г. Ф. Гаузе по экспериментальному изучению конкуренции и хищничества (Гаузе, 1933, 1934а, 19346, 1934в, 1945; Gause, 1932, 1934, 1935). Эти публикации оказали сильное стимулирующее влияние на развитие лабораторных экологических исследований, в результате чего были выполнены всесторонние исследования, представленные работами А. Кромби (Crombie, 1945, 1947), С. Ютида (Utida, 1957), К. Хаффэйкера (Huffaker, 1958), Т. Парка (Park, 1962) и других, что способствовало расширению наших представлений о межпопуляционных взаимодействиях. В то же время многие ученые справедливо поднимали вопрос о том, в какой степени выводы, полученные при лабораторных экспериментах, применимы в полевых условиях к реальным экосистемам. Например, весьма осторожно оценивал значение лабораторных экологических экспериментов Д. Н. Кашкаров, который считал, что экспериментальный метод играет в арсенале экологии второстепенную роль (Кашкаров, 1938). Еще более радикально высказался на эту тему Дж. Хатчисон, сравнивший в шутку лабораторную экспериментальную установку с аналоговой вычислительной машиной, которая дает результат, заранее запрограммированный экспериментатором. Например, при изучении конкуренции двух видов возможно, изменяя условия опыта, добиться как сосуществования обоих видов, так и победы одного из них или, наконец, гибели обоих видов. Подобным же образом в экспериментах по хищничеству или паразитизму, манипулируя условиями культивирования (физические, химические факторы, наличие пли отсутствие убежища для жертвы и т. д.), можно воспроизвести случаи как устойчивого сосуществования в колебательном или стационарном режиме, так и полного выедания жертвы хищником или «ускользания» жертвы из-под контроля хищника. Аргументам Дж. Хатчинсона трудно отказать в убедительности. В дополнение к ним можно привести ряд других доводов, выдвигавшихся различными учеными, которые указывали па необходимость осторожного подхода к оценке значения экспериментов над популяциями в лабораторных условиях для теории природных экосистем (см. Лэк, 1957, 155—156; Макфедьен, 1965, 246—251; Шовен, 1970; Уильямсон, 1975). Принимая это во внимание, нетрудно понять, почему наряду с наблюдением ведущее значение в экологических исследованиях приобрели полевые эксперименты, несмотря на то что в них не может быть обеспечен столь же высокий уровень контроля экспериментатора над изучаемой системой, как в лабораторных условиях. Многочисленные примеры успешного проведения полевых экспериментов при изучении отдельных экологических проблем, таких как влияние метеорологических и эдафических факторов, обеспеченности пищей, конкуренции, хищничества, паразитизма и т. д., известны в сельскохозяйственной науке, лесоведении, охотоведении, ихтиологии, паразитологии и других близких к экологии дисциплинах. Кроме этих тщательно планируемых специально проводившихся опытов немалый вклад в развитие экологической теории был внесен путем обработки и обобщения результатов многочисленных непреднамеренных (стихийных) «экспериментов» с экосистемами, которые были следствием естественных природных процессов или деятельности человека. Среди «экспериментов», поставленных самой природой, результаты которых удалось зарегистрировать, можно указать образование и заселение ряда островов, лавовых полей и других участков terra nullius, изменение гидрологического режима, наступление серии холодных или теплых годов, «удобрение» путем отложения вулканических, аллювиальных или эоловых осадков и т. п. Первыми работами такого рода были комплексные исследования лесных экосистем (бтиогеоценозов), развернувшиеся под руководством В. Н. Сукачева (1956, 1961, 1964), а также радиоэкологические исследования Н. В. Тимофеева-Ресовского (1957, 1961, 1964) и ряда американских ученых (Woodwell, 1962, 1963; Schultz, Klement (еd.) 1963; и др.). В последующие годы происходит широкое развертывание комплексных экспериментальных исследований наземных и водных экосистем в рамках Международной биологической программы (1964-1974) и других научных проектов(Нечаева, Приходько, 1966; Карпов, 1969, 1974; Молчанов, 1970, 1975; Тихомиров, 1971, 1974; Ellenberg (еd.), 1971; Van Dyne, 1972, 1978; Mariscal (еd.), 1974; Likens, Borman, 1975; Van Dупе, Anway, 1976). В итоге, характеризуя значение экспериментального метода в экологии, необходимо признать, что его применение, особенно на основе достижений научно-технического прогресса, открывает перед экологией грандиозные перспективы. Однако экологический эксперимент, равно как и наблюдения над экосистемами, становится наиболее эффективным только в сочетании с третьим важнейшим методом экологии - методом моделирования.
Моделирование
Весь опыт развития естествознания вообще и гидроэкологии в частности свидетельствует, что наблюдения и эксперименты в наибольшей степени способствуют познанию лишь тогда, когда они задуманы и осуществлены на основе научной теории. В точных естественных науках, к каковым все более стремится и современная экология, весьма эффективной формой выражения теоретических представлений выступают модели, а одним из наиболее плодотворных методов служит метод моделирования, т. е. построения, проверки, исследования моделей и интерпретации полученных с их помощью результатов.
|
|||||||||||
|
|
Последнее изменение этой страницы: 2016-08-26; просмотров: 488; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.224.214.215 (0.046 с.) |