Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Ядро нейрона, цитоплазма, вещество Ниссля, нейрофибриллы, митохондрии и другие включения
Ядро нервной клетки отличается сравнительно большими размерами, круглой или овальной формой. Объемное соотношение между ядром и цитоплазмой клетки значительно варьирует в различных образованиях нервной системы. Мелкие клетки обычно имеют относительно более крупное ядро. Ядро нервной клетки содержит ядерный сок (кариоплазму), в котором различными гистологическими и гистохимическими методами выявляются гранулы, содержащие рибонуклеопротеид (хроматин). Оболочка ядра сравнительна плотна и под электронным микроскопом выявляется в виде двойной мембраны с нерегулярно расположенными порами. В некоторых нервных клетках оболочка состоит из нескольких мембран, заметно варьирующих по диаметру. Внутри ядра, кроме гранул кариоплазмы, находится ядрышко, диаметр которого достигает 0,5-1 мкм. Ядрышко содержит рибонуклеиновую (РНК) и дезоксирибонуклеиновую (ДНК) кислоты. На электронно-микроскопических снимках вещество ядрышка имеет вид сложной ячеистой сети без какой-либо наружной мембраны. Некоторые исследователи наблюдали тесную связь между веществом ядрышка и кариоплазмой. Показано также, что между кариоплазмой и цитоплазмой нервной клетки происходит обмен нуклеопротеидами. В ядрах двигательных клеток ДНК располагается в виде мелких зёрнышек, диффузно распределенных по всей массе кариоплазмы. Наряду с этим имеются от двух до десяти зёрнышек, дающих более интенсивную окраску на ДНК, и несколько перинуклеарных зёрнышек. В ядрах чувствительных нейронов спинномозговых узлов зерна хроматина расположены более рыхло и выделяются только 2-3 перинуклеарных зёрнышка. В ядрах центральных переключательных нейронов ДНК содержится в малых количествах. Таким образом, ядро нервной клетки является центральным мембранным элементом нейрона, содержащим прозрачный ядерный сок (кариоплазму), в котором находятся рибонуклеопротеид и ядрышко, содержащее рибонуклеиновую и дезоксирибонуклеиновую кислоты. Гистохимические и электронно-микроскопические исследования показали, что структура органоидов определяется их химическим составом и функциональным значением в комплексных обменных процессах. Вещество Ниссля (тигроид) является специфическим органоидом нервной клетки. Тигроид на препаратах, окрашенных основными красителями (метиленовый синий, тионин, галлоцианин, крезилвиолетт, пиронин), наблюдается в виде хромофильной зернистости с определённой ориентацией. Тигроид обнаруживается прижизненно с помощью фазово-контрастной микроскопии. Распределение тигроида в нервной клетке имеет ряд характерных особенностей в различных отделах нервной системы. Большие различия по данному признаку имеются, например, между клетками корковых зон зрительного и двигательного анализаторов. Л. Ольшевскому удалось на основе анализа топографии и содержания тигроида в нервной клетке выделить некоторые трудно дифференцируемые ядра ретикулярной формации. Как правило, в крупных нейронах тигроид содержится в больших количествах и более равномерно распределён внутри тела клетки, в отличие от нервной клетки меньших размеров. Глыбки тигроида проникают и в отходящие от клетки дендриты. Гистохимические исследования показали, что тигроид является нуклеопротеидом. Исследование тигроида способом замораживания и высушивания под вакуумом выявило, что глыбки распределяются в цитоплазме нервной клетки неравномерно. Исследования с рибонуклеазой обнаружили, что в состав нисслевской субстанции входит РНК. Обработка срезов этим ферментом делает глыбки невосприимчивыми к окраске основными красителями. Однако основа глыбок при этом сохраняется. В их состав входит по крайней мере два вида белков: один из них имеет щелочную реакцию и богат аргинином и гистидином, другой же обладает нейтральной или кислой реакцией и содержит ароматические аминокислоты.
В онтогенезе появление тигроида в дифференцированной форме совпадает по времени с включением нейрона в функциональную систему и поэтому происходит в различных отделах нервной системы в разные сроки. Нейрофибриллы являются вторым важным компонентом цитоплазмы нервной клетки и хорошо выявляются различными методами: импрегнацией серебром, а также прижизненной окраской метиленовым синим. Они были обнаружены в нервной клетке не только позвоночных, но и беспозвоночных. Под микроскопом они представляют собой тонкую сеть, в петлях которой лежат зёрна тигроида. При рассмотрении в электронный микроскоп в теле нервной клетки, как правило, можно выявить микротрубочки (диаметром 20-30 нм), нейрофиламенты (10 нм) и микрофиламенты (5 нм). В основе химической структуры нейрофибрилл лежат белки. Наличие микротрубочек и нейрофиламентов в аксонах и дендритах позволило предположить, что они участвуют в транспортировке различных веществ, что находит подтверждение в ряде биохимических исследований. Микрофиламенты изобилуют в растущих нервных отростках, их много также в нейроглии. Полагают, что микрофиламенты принимают участие в управлении движением клеточной мембраны и подлежащей цитоплазмы. Некоторые авторы считают также, что нейрофибриллы являются основным субстратом для проведения нервных возбуждений с участием в этом процессе мембранных поверхностей тела клетки и её отростков, другие же относят нейрофибриллы к опорному аппарату. Специфичность нейрофибрилл для нервной клетки подвергается некоторыми исследователями сомнению, поскольку подобные нити обнаружены на электроннограммах клеток самых разнообразных тканей.
Митохондрии нервной клетки имеют зернистую, палочковидную или нитевидную форму. Их количество варьирует в теле нервной клетки в очень больших пределах. Особенно много их на месте выхода аксона из нервной клетки (аксонный конус или холмик) и в области синапсов. Гистохимические исследования показали, что в митохондриях нервной клетки имеются рибонуклеопротеиды, содержание которых меняется в зависимости от состояния клетки. В них установлена локализация различных ферментных систем, в частности, окислительных ферментов – сукцинатдегидрогеназа и цитохромоксидазы, активность которых возрастает во время возбуждения нервной клетки и падает при её утомлении. Наряду с этими ферментами в митохондриях нервной клетки обнаруживаются также некоторые неспецифические ферменты, например, кислая фосфатаза. Под электронным микроскопом внутренняя структура представляется весьма сложной. Каждая митохондрия имеет наружную двойную мембрану, от которой в просвет митохондрии отходит ряд гребешков, делящих её полость на отсеки. В последних работах показано, что эти гребешки (кристы) являются местом локализации названных выше ферментных систем. Митохондрии нервной клетки резко меняют свою форму в зависимости от того, локализованы ли они в теле или в её отростках. В теле клетки кристы митохондрии имеют перпендикулярное или косое расположение относительно длинной их оси; в дендритах же кристы располагаются параллельно длинной оси митохондрии. В аксонах митохондрии достигают необычайной длины (до 10 микрон) и иногда имеют сложную ветвистую форму. Включения. В нервной клетке часто обнаруживаются пигментные гранулы. Тёмно-коричневые или чёрные гранулы меланина постоянно встречаются в нервной клетке некоторых отделов ЦНС (средний мозг – чёрная субстанция, продолговатый мозг – locus coeruleus, дорсальное ядро блуждающего нерва, а также межпозвоночные и симпатические узлы). Значение этих пигментных образований пока неясно. В нервной клетке часто встречаются также желтые гранулы липофусцина, являющегося продуктом нормальной жизнедеятельности нервной клетки. Количество липофусцина увеличивается с возрастом. Накопление жира в нервной клетке может являться результатом как нормального, так и патологического обмена. Гликоген обнаруживается во многих нервных клетках. Показано, что в цитоплазме ряда крупных нейронов, в особенности двигательных, содержится гликоген, количество которого меняется в зависимости от функционального состояния нервной клетки. По некоторым данным, гликоген связан с тигроидом. Гликоген обнаружен также в нервных окончаниях в области синапсов.
Железосодержащие гранулы выявлены в нервной клетке чёрной субстанции, бледного шара и некоторых других образований ЦНС. С возрастом количество железосодержащих гранул возрастает. При прижизненном воздействии на нервные клетки основных или кислых красителей происходит, как и в других клетках организма, их отложение в виде гранул в области аппарата Гольджи, что можно рассматривать как защитное приспособление нервной клетки.
Размножение клеток
С синтезом веществ протоплазмы и ростом теснейшим образом связано размножение клеток, осуществляющееся путём деления. Основная и универсальная форма деления клеток – митоз (непрямое деление, или кариокинез). При митозе происходит сложная реорганизация ядра и цитоплазмы, которая в наиболее полной форме приводит к образованию двух дочерних клеток, подобных исходной материнской. В процессе митотического деления различают 4 основные стадии: профазу, метафазу (стадия материнской звезды), анафазу (стадия дочерних звёзд) и телофазу. К началу деления животные клетки обычно округляются; центриоли начинают отдаляться друг от друга; цитоплазма между ними приобретает фибриллярное строение; вокруг центриолей путём образования радиально расположенных нитчатых структур образуется лучистое сияние – астросфера. Вскоре центриоли оказываются у двух противоположных полюсов клетки, а фибриллярно структурированная цитоплазма вытягивается между ними в ахроматиновое веретено – расходящиеся микротрубочки клетки. Сущность митоза состоит в преобразовании ядерного аппарата. При переходе интеркинетичекого ядра в стадию профазы в нём вместо глыбок хроматинового материала становятся видимыми микроскопические нитчатые образования – хромосомы. Во многих случаях на этой стадии по длине хромосом обнаруживаются варикозные, интенсивно окрашивающиеся утолщения – хромомеры, имеющие в каждой хромосоме определённую форму и расположение.
В течение профазы происходит укорочение хромосом до 1/10-1/20 первоначальной длины при одновременном их утолщении – спирализация хромосом – результат спирального скручивания входящих в состав хромосом тончайших нитей – хромонем. После спирализации хромомерная структура становится незаметной, и хромосомы превращаются в равномерно окрашивающиеся тела, состоящие из двух продольно сложенных, но не всегда различимых половин – хроматид. В это же время исчезает оболочка ядра и растворяется ядрышко. На стадии метафазы хромосомы располагаются в экваториальной плоскости клетки, образуя фигуру материнской звезды или экваториальную пластинку; на этой стадии особенно легко определяются число и форма хромосом. Установлено, что во всех делящихся соматических клетках данного животного организма или спорофита у растений с чередованием поколений, как правило, имеется двойной – диплоидный набор различных по форме и величине хромосом, в зрелых половых клетках хромосом – одинарный (гаплоидный набор или геном). Диплоидный набор хромосом является результатом соединения при оплодотворении хромосомных комплексов женской и мужской половых клеток. Таким образом, в диплоидном наборе каждый сорт хромосом представлен двумя гомологичными хромосомами материнского и отцовского происхождения. У разных видов количество хромосом сильно варьирует. Минимальное количество хромосом у аскариды – 1 пара; у ракообразных имеется 200 хромосом. В тканях человека насчитывается, по данным одних авторов, 23, других – 24 пары хромосом. Важнейшей структурной частью каждой хромосомы является маленький слабо окрашивающийся участок – центромера, или кинетическое тельце. При расщеплении хромосомы на 2 хроматиды этот участок также раздваивается. При достижения хромосомами экваториальной плоскости веретена центромера оказывается местом прикрепления нитей, идущих от полюса веретена, – хромосомных нитей. При этом к центромере одной хроматиды прикрепляется хромосомная нить от одного полюса, а к центромере другой хроматиды той же хромосомы – нить от противоположного полюса. Хромосомные нити располагаются на переферии веретена, остальные же его нити идут от полюса к полюсу клетки, не прерываясь. В стадии анафазы хроматиды каждой хромосомы начинают расходиться к своим полюсам; образуются две группы направляющихся к полюсам хромосом – дочерние звёзды. На последней стадии – телофазе – происходит реконструкция интеркинетического ядра. Хромосомы в результате деспирализации вновь удлиняются и утончаются, становясь плохо различимыми. Определённые участки хромосом остаются в интеркинетическом ядре спирализированными и удерживают большие количества ДНК – они имеют вид глыбок гетерохроматина. Кариосомы представляют собой крупные скопления гетерохроматина. Деспирализующиеся участки хромосом – эухроматиновые участки – бледно окрашиваются и становятся плохо различимыми. В телофазе восстанавливается ядерная оболочка и ядрышко. Способ возникновения этих структур неясен; предполагают, что в формировании ядрышек участвуют особые, так называемые ядрышковые, хромосомы. Веретено и астросфера претерпевают обратное развитие. По экватору клетки образуется кольцевая перетяжка, приводящая к отделению дочерних клеток друг от друга (плазматомия).
В процессе митоза митохондрии и аппарат Гольджи более или менее равномерно распределяются между дочерними клетками. Если аппарат Гольджи имеет компактную форму, то в начале деления он предварительно распадается на отдельности (диктиосимы), из которых в дочерних клетках вновь реконструируется исходная структура аппарата (диктиокинез). В период митоза у дифференцированных клеток выполнение специализированных функций, например, выработка секрета, обычно прерывается. Могут на это время исчезать, а затем восстанавливаться некоторые специальные структуры и включения, например, реснички в мерцательных клетках, гранулы секрета в железистых. В клетках высших растений митоз протекает без участия центросом и без образования астросфер (анастральный тип); различные отклонения от описанной схемы могут наблюдаться у простейших (например, у инфузорий при делении микронуклеуса ахроматиновое веретено располагается внутри ядерной оболочки), однако сущность митотического деления – это совершенно точное распределение хромосомного материала между дочерними ядрами путём продольного расщепления хромосом и последующего расхождения их половин к разным полюсам материнской клетки – во всех случаях одинаков. Значение митоза и смысл его общебиологического распространения могут быть поняты в настоящее время только на основе хромосомной теории наследственности, согласно которой хромосомы являются носителями факторов наследственности – генов, каждый из которых принимает специфическое участие в изменении веществ в клетке. Для нормального развития и функционирования организма необходим полный набор генов, а следовательно и полный набор хромосом, так как каждая хромосома содержит лишь определённую часть генов данного организма. Гены расположены в линейном порядке вдоль оси хромосомы; гомологичные хромосомы имеют ассортимент генов одинокого назначения. Ряд фактов заставляет предположить, что специфичность каждого гена определяется специфичностью входящих в состав хромосомы молекул ДНК (и их частей), связанных с основным белком. Молекулы ДНК состоят из двух спирально закрученных нитей (Уотсон и Крик); каждая нить представляет собой высокополимерное соединение, построенное из множества элементарных частиц ДНК – нуклеотидов; имеется 4 вида нуклеотидов, которые различаются структурой своих азотистых оснований; специфичность молекулы ДНК обусловлена своеобразием в чередовании различных нулеотидов. Размножение клеток предполагает умножение генного материала и равное распределение его между дочерними клетками. Первая осуществляется путем построения – редупликации новых аналогичных генных макромолекул. Редупликация, по-видимому, происходит в конце интеркинеза. Точное распределение генов между клетками достигается митозом благодаря продольному расщеплению каждой хромосомы на две одинаковые дочерние хроматиды. Особой формой митоза является деление, связанное с образованием половых клеток – мейоз. У всех многоклеточных животных, ряда простейших и у низших растений мейоз непосредственно предшествует образованию половых клеток. У остальных организмов он может осуществляться на других фазах жизненного цикла. Мейоз состоит из 2-х последовательно протекающих делений, в результате которых из диплоидных ооцитов и сперматоцитов 1-ого порядка образуются яйцевые клетки и сперматозоиды с гаплоидным набором хромосом. Достигается это тем, что в профазе первого мейотического деления происходит сближение – конъюгация гомологичных хромосом, а затем в анафазе первого мейотического деления к каждому полюсу отходят по одной целой дуплицированной хромосоме от каждой гомологичной пары. В анафазе 2-ого мейотического деления к полюсам расходятся хроматиды от каждой дуплицированной хромосомы. Таким образом, переход диплоидного набора хромосом к гаплоидному осуществляется уже при первом мейотическосм делении (в некоторых случаях расхождение гомологичных хромосом происходит при 2-м мейотическом делении). При оплодотворении восстанавливается диплоидный набор хромосом. Значение мейоза не ограничивается предотвращением удвоений хромосом в каждом следующем поколении. При конъюгации хромосом во время первого мейотического деления гомологичные хромосомы обмениваются отдельными своими участками. Это приводит к перекомбинации части отцовских и материнских генов в хромосоме и таким образом повышает наследственную изменчивость в следующих поколениях. В тканевых клетках митоз может не завершаться плазматомией; это приводит к образованию двуядерных клеток. Иногда в клетке умножение набора хромосом осуществляется без увеличения числа ядер – эндомитоз. При эндомитозе не происходит реорганизации цитоплазмы, сохраняется ядерная оболочка и ядрышко; размеры ядра увеличиваются. Эндомитоз наблюдается в различных тканевых клетках животных и растений. Большое распространение он имеет у простейших, где приводит, в частности, к образованию крупных макронуклеусов инфузорий, заключающих в себе большое число хромосомных наборов. Ядра с увеличенным числом хромосомных наборов называются полиплоидными; имеющие четыре набора вместо двух называются тетраплоидными, восемь – октаплоидными и т.д. Полиплоидность может быть получена не только путем эндомитоза, но и в результате повреждения механизма митоза при ряде воздействий на делящиеся клетки (например, при отравлении колхицином, аценафтеном, трипафлавином и др.). Полиплоидность связана с увеличением размеров клеток – при полиплоидности всех тканевых клеток обычно увеличиваются и размеры организма. В некоторых тканевых клетках (например, у насекомых) происходит умножение хромонем в каждой хромосоме без увеличения числа хромосом и без характерной для митоза реорганизации ядра и цитоплазмы – политения. При этом хромосомы становятся крупнее и по размеру могут превышать в сотни раз обычные хромосомы (например, во многих соматических клетках двукрылых). Политения также сопровождается увеличением размеров клетки. При политении и полиплоидии в ядрах соответственно возрастает количество ДНК. Эндомитоз и политения в специализированных или дифференцирующихся клетках приводят к увеличению ядерного материала без нарушения их специфических функций и структур. В клетках различных нормальных (скелетные мышцы, фиброциты, эпителий мочевого пузыря и т.д.) и патологических (злокачественные опухоли) тканей можно наблюдать деление ядер путем амитоза. Нередко амитозы наступают после повреждения тканей разными агентами. При амитозе ни в ядре, ни в цитоплазме не обнаруживаются какие-либо существенные перестройки. Амитоз обычно начинается с перешнуровывания ядрышек, затем перетяжкой или путем образования перегородки ядро разделяется на две половинки. Иногда ядро сразу делится на несколько частей (фрагментация). В некоторых случаях деление происходит неравномерно, и от ядра отпочковываются небольшие части кариоплазмы. Обычно амитоз не сопровождается плазматомией и приводит лишь к увеличению числа ядер в клетке. Вопросы полноценности амитоза как способа деления ядер, состояния хромосомного аппарата при амитозе и возможности смены амитоза митозом' до настоящего времени не решены. Материнский и отцовский организмы одинаково влияют на наследственность потомков, несмотря на то, что в оплодотворенном яйце – зиготе цитоплазма почти полностью материнского происхождения. Это обстоятельство свидетельствует о том, что основными носителями наследственности служат хромосомы ядра, которые содержатся в яйцевой клетке и сперматозоиде в равном количестве (не считая половых хромосом). Наряду со многими другими фактами этот вывод подтверждается прямыми опытами, в которых зигота, содержащая цитоплазму яйца одного вида и ядро сперматозоида другого, развивалась в организм, обладавший только признаками отца (опыты на шелковичных червях Б.Л. Астаурова). Имеются, однако, данные, свидетельствующие о том, что некоторые факторы наследственности находятся и в цитоплазме. На это указывают, в частности, те случаи, когда признак передается матерью, но не передается отцом. Явления цитоплазматической наследственности обнаружены в значительном количестве у высших и низших растений, в меньшем – у животных (например, пестролистность у некоторых декоративных растений, вязкость цитоплазмы, устойчивость к ядам, температурный оптимум и др. свойства у кипрея, чувствительность к CO2 у дрозофилы и т.д.). Факторы наследственности должны быть способны к самовоспроизведению. В цитоплазме животных клеток среди микроскопически видимых структур к редупликации путем деления способны митохондрии, аппарат Гольджи и центриоли, у растительных клеток – митохондрии и пластиды. Однако не исключена также возможность образования этих структур в цитоплазме заново, что было неоднократно показано в отношении центриолей. Вопрос о том, какие компоненты цитоплазмы могут быть носителями факторов наследственности, остается открытым; лишь в некоторых случаях эту роль с большой долей вероятности удается приписать хлоропластам. Одной из важнейших, но не решенных проблем цитологии является выяснение того, каким путем хромосомы оказывают специфическое влияние на обмен веществ клетки и тем самым определяют их наследственно обусловленные свойства. Для решения этого вопроса и определения участия ядра в осуществлении клеточных функций большое значение имеют наблюдения над клетками, искусственно лишенными ядра, а также опыты по введению в клетку с удаленным ядром ядра, взятого от клетки другого вида (Хеммерлинг). Синапс нервной клетки
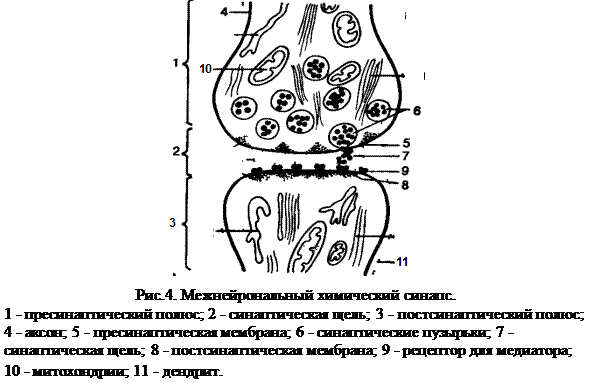
Синапс – (от греческого synapsis - соприкосновение, связь) - место контакта двух нейронов или нейрона и мышцы.
Синапс состоит из 2-х мембран, соприкасающихся друг с другом: одна из них принадлежит разветвлению аксона одного нейрона, а другая – дендриту другого нейрона (рис. 4.). При исследовании синапса под электронным микроскопом ясно видна граница контактирующих друг с другом нейронов. На этой границе чётко вырисовываются две мембраны – пресинаптическая и постсинаптическая, отделённые друг от друга синаптической щелью. В центральной нервной системе синаптическая щель является непосредственным продолжением межклеточного пространства, их содержимое сообщается друг с другом. Ширина синаптической щели – от 2 до 30 нм, диаметр синаптического контакта – от 0,1 до 10 мкм. Синаптическая щель – промежуток, разделяющий пресинаптическую мембрану аксона одной клетки и постсинаптическую мембрану тела или дендрита нейрона другой клетки или мышцы. Пресинаптическая мембрана является продолжением поверхностной мембраны аксонального окончания, глиальные элементы не участвуют в образовании синапсов. Эта мембрана не сплошная, она имеет отверстия, через которые цитоплазма аксональных окончаний сообщается с синаптическим пространством. Постсинаптическая мембрана менее плотная, чем пресинаптическая, она не имеет отверстий. Толщина каждой из мембран синапса не превышает 5-6 нм. Несколько иначе построены органные синапсы, например, в области нервно-мышечного соединения. На поверхности мышечного волокна имеется углубление со множеством ветвящихся и взаимодействующих между собой складок, в которых размещаются разветвления аксона. Здесь также различаются пресинаптическая (аксональная) и постсинаптическая (мышечная) мембраны. Обе мембраны состоят из нескольких слоев, толщина каждой – около 10 нм; пространство между мембранами заполнено сильно гидратированным гелем. Синапсы бывают двух видов – возбудительные и тормозные, с их помощью происходит соответственно передача или блокада нервного импульса. Основной функцией синапса является передача возбуждения с одной нервной клетки на другую, либо с нейрона на эффекторный орган. По современным данным, в большинстве синапсов передача возбуждения осуществляется посредством медиатора, синтезируемого и накапливаемого в нервных окончаниях. Об этих функциях будет подробно рассказано при изучении физиологии ЦНС.
|
||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 1371; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.129.19.251 (0.035 с.) |