Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Эпигенетическая теория эволюции ⇐ ПредыдущаяСтр 10 из 10
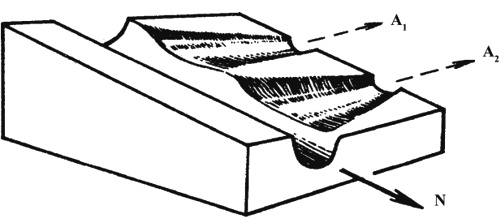
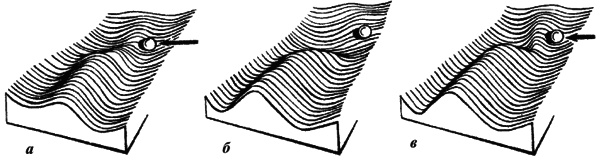
Основные положения В отличие от синтетической, эпигенетическая теория видит эволюционный процесс прежде всего как процесс эволюционного преобразования онтогенеза (Шишкин, 1987, 1988а, б, 2006; Раутиан, 1993). При этом особое внимание обращается на целостность онтогенеза в том смысле, что и сам процесс, и его результат (строение организма на последовательных этапах его развития) гораздо устойчивее, чем любые отдельные факторы и процессы развития. Нормальное развитие эквифинально и способно релаксировать (подавлять, поглощать, нивелировать) очень широкий круг воздействий и возмущений, как внешних, так и внутренних, включая результаты различных ошибок и нарушений нормального развития. Таким образом, наследуется (устойчиво воспроизводится в последующих поколениях) нормальный онтогенез (норма реакции) в целом, а не отдельные признаки. Помимо нормы, существует масса разнообразных уклонений (аберраций) развития с неустойчивым воспроизведением, которые в нормальных условиях реализуются редко. В неблагоприятных условиях, когда механизмы защиты нормы удается нарушить или преодолеть — в результате ли сильного внешнего воздействия или по внутренним причинам (ошибки развития), — развитие идет по аберрантному (уклоняющемуся и неустойчивому) пути. Поскольку эндогенные аберрации развития легко возникают не только при половом, но и при партеногенетическом и бесполом размножении, их возникновение есть реализация скрытой гетерогенности популяции, а не возникновение de novo каких-то генетических изменений. Эпигенетическая теория рассматривает аберрации как материал отбора, который способен создать на ее основе новую адаптивную норму. Такой процесс легче описать в рамках метафоры эпигенетического ландшафта. Нормальный онтогенез представляет собой устойчивую (стабилизированную) последовательность тесно связанных отдельных эпигенетических процессов. Более или менее обособленные отрезки этой последовательности именуют креодами. Если представить себе креод в виде сильно заглубленной долины в некотором ландшафте, по уклону которой течет онтогенетический процесс (рис. 1), то аберрациями будут пологие боковые долинки, приподнятые в бортах главной. Чтобы аберрация реализовалась, т.е. чтобы онтогенез мог выйти в боковую долину, нужно либо сильное внешнее воздействие на развивающийся организм, выталкивающее его на боковую долину (рис. 2а), либо изменение самого ландшафта, выполаживающее дно долины (дополнительное аберративное изменение, рис. 2б), либо то и другое вместе (рис. 2в).
Рис. 1. Участок эпигенетического ландшафта, показывающий перепады высоты стенок (порогов устойчивости) креода в местах ответвления аберративных долин. А1, А2 - аберративные долины, N - главная долина (креод) (по Шишкину, 1987) Аберрации мало устойчивы («не наследственны»; более устойчивые из них часто именуются модификациями), но они постоянно, хотя и непредсказуемо в деталях, воспроизводятся в популяции, поскольку эпигенетический ландшафт с соответствующими боковыми долинками — это адаптивная норма вида. Поэтому, несмотря на свою слабую наследуемость, именно аберрации служат материалом отбора. Если аберрация оказалась полезной, отбор будет избирательно сохранять те эпигенотипы, которые более устойчиво ее воспроизводят. Другими словами, преимущество получат те эпигенотипы, где соответствующая долинка углублена, и уровень ее дна на стыке с главной долиной более приближен ко дну последней. Если при этом еще и главная долина будет выполаживаться ниже развилка (т.е. если понизится устойчивость прежней нормы), прежняя аберрация будет иметь шанс стать новой нормой. Если же прежняя норма сохранит свое значение, то стабилизируются обе нормы и будет выработан механизм онтогенетического переключения между ними. В случае когда этот механизм использует для переключения средовой сигнал, мы получаем типичную модификационную изменчивость. Там, где используется генетический сигнал (например, рекомбинация), мы получим одну из форм менделирования. Таким путем менделирующие признаки возникают и в природе (например, обычные механизмы определения пола), и в лаборатории (например, в процессе стабилизации чистых линий).
Рис. 2. Соотношения между строением эпигенетического ландшафта и характером повреждающего воздействия: а - уклонение развития на боковую долину за счет сильного внешнего воздействия (длинная стрелка); б- такое же уклонение под действием сильной мутации, вызывающей нарушение креода; в - промежуточное состояние (по Шишкину, 1987)
В целом для эпигенетической теории эволюции зафиксированный («запомненный») в адаптивной норме спектр возможных путей развития (креодов и их аберраций), т.е. эпигенетический ландшафт — это сущность живого организма, его онтогенетический потенциал, то, что определяет существование организма и с чем работает отбор. Чисто генетический уровень (гены и их аллели, динамика их частот в популяции, мутации, рекомбинация и т.п.) лежит гораздо глубже и не определяет специфики эволюционных процессов. Точно так же из процессов и закономерностей квантово-механического уровня нельзя вывести специфики того, что происходит на макроуровне. «Гены приходят и уходят, а креоды остаются»: хорошо известно, что признаки более устойчивы, чем гены и аллели, будто бы их определяющие. Более подробно онтогенетические процессы в аспекте наследования рассмотрены М.А. Шишкиным (1987, 1988а, 2006). Для нас же более важна другая сторона проблемы эволюции онтогенеза.
|
||||||
|
|
Последнее изменение этой страницы: 2016-08-15; просмотров: 631; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.139.97.157 (0.006 с.) |