Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Общая характеристика источниковСодержание книги Поиск на нашем сайте
ОБЩАЯ ХАРАКТЕРИСТИКА ИСТОЧНИКОВ И ПУТЕЙ РАСХОДОВАНИЯ АМИНОКИСЛОТ В ТКАНЯХ
Совокупность свободных аминокислот в живых организмах образует так называемый аминокислотный фонд. У взрослого человека этот фонд составляет примерно 35 г. Обычно процессы расходования и пополнения аминокислотного фонда уравновешены, то есть он находится в состоянии динамического равновесия. Пополнение аминокислотного фонда осуществляется за счет эндогенных и экзогенных источников. За счет эндогенных источников пополняется 2/3, а за счет экзогенных – 1/3 аминокислотного фонда. Одним из эндогенных источников аминокислот в организме является синтез заменимых аминокислот. Из 20-ти входящих в состав белков аминокислот у человека могут синтезироваться 10 (табл.1). Остальные 10 аминокислот должны поступать с пищевым белком. Следует отметить, что такие аминокислоты, как аргинин и гистидин, являются частично заменимыми, поскольку они синтезируются в организме, однако со скоростью, не достаточной для того, чтобы обеспечить рост организма в детском возрасте. Таблица 1
Вторым эндогенным источником пополнения аминокислотного фонда является распад белков организма. У здорового человека обновление белков составляет 1–2% от общего количества белков тела, или 30–40 г в сутки. При этом примерно 75–80% высвободившихся аминокислот снова используется в синтезе белков. Оставшаяся часть подвергается катаболизму до конечных продуктов азотистого обмена, превращается в глюкозу, кетоновые тела или жирные кислоты. Поступление аминокислот в составе пищевого белка является экзогенным источником пополнения амикислотного фонда. Для поддержания стационарного состояния аминокислотного фонда взрослый организм нуждается в потреблении примерно 60 г белка. При этом важным является качество пищевого белка, которое оценивается по наличию незаменимых аминокислот в пищевом белке и соответствию их соотношения в синтезируемых организмом белках. С учетом аминокислотного скора (наличия и соотношения незаменимых аминокислот), а также перевариваемости и усвояемости, пищевые белки можно расположить по ценности в следующем порядке: яичный белок, альбумин молока, белки рыбы, мяса, белки растительных продуктов. Недостаточное содержание в рационе полноценных белков приводит к развитию кахексии и квашиоркора (детская дистрофия). Расходование аминокислотного фонда связано со следующими процессами: 1. Синтез белков и пептидов. 2. Синтез соединений других классов: а) пуриновых и пиримидиновых оснований (Гли, Глн, Асп); б) катехоламинов и йодтиронинов (Тир); в) биогенных аминов (Глу, Гис, 5-гидрокситриптофан); г) порфиринов (Гли); д) глицерофосфолипидов (Сер, Мет); е) креатина (Арг, Мет); ж) меланинов (Тир, Цис); з) витамина РР (Три). 3. Распад ради энергии. Около 10% от общего количества АТФ при смешанной диете образуется за счет окисления аминокислот. 4. Образование глюкозы (все аминокислоты, за исключением Лиз и Лей). 5. Образование кетоновых тел и жирных кислот (Лей, Йле, Лиз, Тир, Фен). ПЕРЕВАРИВАНИЕ БЕЛКОВ Поскольку белки организма отличаются видовой и тканевой специфичностью, живой организм способен использовать вводимый с пищей белок только после полного его расщепления в ЖКТ до аминокислот, из которых в клетках синтезируются специфические для данного вида белки. Процесс высвобождения аминокислот из белков катализируется специальными ферментами, получившими название протеиназы. Протеиназы – это ферменты, которые относятся к классу гидролаз. По характеру действия их разделяют на эндопептидазы и экзопептидазы. Эндопептидазы разрывают пептидные связи внутри пептидной цепи. К ним относятся: пепсин и гастриксин желудка; трипсин, химотрипсин, эластаза поджелудочной железы. Экзопептидазы катализируют разрыв концевой пептидной связи с отделением концевой аминокислоты. К ним относятся карбоксипептидазы А и В поджелудочной железы, аминопептидазы и дипептидазы тонкого кишечника (мембраносвязанные ферменты). Ферменты, участвующие в переваривании белков, обладают относительной субстратной специфичностью, которая обусловлена тем, что пептидазы быстрее гидролизуют пептидные связи между определенными аминокислотами, что позволяет за более короткое время расщепить белковую молекулу (табл. 2). Таблица 2 Характеристика протеолитических ферментов Желудочно-кишечного тракта
Примечание: Х – любая аминокислота.
Начальные стадии гидролиза белков осуществляются за счет полостного пищеварения эндопептидазами желудочного и поджелудочного соков. В результате полипептиды расщепляются до полипептидов меньшего размера. Главным механизмом промежуточных и заключительных стадий гидролиза является мембранное пищеварение. Оно осуществляется адсорбированными ферментами поджелудочного сока и кишечными мембраносвязанными ферментами. У детей грудного возраста в желудке находится фермент реннин, который отщепляет от основного белка молока казеина гликопротеин, в результате чего образуется параказеин. Параказеин присоединяет ионы кальция, что приводит к створаживанию молока и задержке его выхода из желудка в кишечник. Это создает условия для проявления протеолитической активности пепсина. У взрослых людей створаживание молока происходит под действием соляной кислоты и пепсина. Роль соляной кислоты в переваривании белков
1. Вызывает денатурацию и набухание белков, способствуя тем самым лучшему их гидролизу. 2. Создает рН-оптимум для пепсина (1,5–2,5) и гастриксина (3,5). 3. Осуществляет частичный протеолиз пепсиногена и переводит его в пепсин, т.е. является активатором пепсина. 4. Способствует секреции секретина – гормона пищеварения. 5. Обладает антимикробным действием. От денатурирующего влияния соляной кислоты и переваривающего действия пепсина собственные белки стенок желудка предохраняет слизистый секрет, содержащий гликопротеины.
НОРМАЛЬНЫЕ СОСТАВНЫЕ ЧАСТИ ЖЕЛУДОЧНОГО СОКА В состав нормального желудочного сока входят: вода, белки, пепсин, гастриксин, липаза, муцин, соляная кислота, хлорид натрия, кислые фосфаты, гликопротеин – так называемый внутренний фактор Кастла и другие вещества. Кислотность желудочного сока формируется за счет следующих компонентов: свободная соляная кислота, связанная соляная кислота, кислореагирующие фосфаты, органические кислоты. Соляную кислоту, находящуюся в солеобразном соединении с белками и продуктами их гидролиза, называют связанной соляной кислотой. Несвязанную с белками кислоту называют свободной соляной кислотой. У здорового взрослого человека содержание свободной соляной кислоты составляет 20–40 ммоль/л. Сумма всех кислореагирующих веществ желудочного сока составляет общую кислотность желудочного сока, которая у здорового взрослого человека составляет 40–60 ммоль/л. Одним из основных методов исследования желудочной секреции является зондовый, с его помощью производится отбор проб желудочного сока с последующим исследованием его компонентов и состояния кислотности. Этот метод является нефизиологичным и травматичным, в связи с чем противопоказан некоторым людям (сужение пищевода, нарушения психики). Существуют и беззондовые методы, основанные на использовании ионообменных смол, способных обменивать ионы в кислой среде. При применении этого метода кислотность желудочного сока оценивается по изменению цвета мочи. Кроме того, используется десмоидная проба, которая основана на использовании резинового мешочка с метиленовым синим, перевязанного кетгутовой нитью № 3. Этот мешочек заглатывается. Если кислотность желудочного сока нормальная, кетгутовая нить перева-ривается под действием пепсина, метиленовый синий всасывается. Этот факт устанавливается по изменению цвета мочи. Если моча сохраняет обычный цвет, кислотность желудочного сока понижена или в нем отсутствует соляная кислота. Определение кислотности желудочного сока важно для диагностики различных заболеваний желудка. Повышенная кислотность желудочного сока сопровождается изжогой, диареей и может быть симптомом язвенной болезни желудка и двенадцатиперстной кишки, а также гиперацидного гастрита. Пониженная кислотность или полное отсутствие соляной кислоты наблюдается при атрофических гастритах и часто сопровождается анемией вследствие недостаточной выработки фактора Кастла. Анацидность, или ахлоргидрия, свидетельствует о значительной потере слизистой оболочкой обкладочных клеток, что может быть при раке желудка. Желудочный сок ребенка до года характеризуется невысокой кислотностью (рН 3,0–4,0, в единицах кислотности). Причем до 4–5-месячного возраста кислотность обусловлена присутствием молочной кислоты, а позже и соляной. Интенсивность секреции желудочного сока возрастает примерно в два раза при смешанном вскармливании. Первые 2 месяца жизни главную роль в переваривании белков играет ренин, а затем гастриксин и пепсин. Однако первые два года активность протеиназ остается низкой и достигает максимального уровня к 3-м годам. Ферментативная активность слизистой кишечника высокая, преобладает мембранное пищеварение. Женское молоко в желудке находится 2–3 часа, питательная смесь с коровьим молоком – 3–4 часа. Белки женского молока перевариваются и всасываются лучше, чем коровьего, и усваиваются на 90–95%, а коровьего – на 60–70%. Рис. 1. g-глутамильный цикл всасывания аминокислот
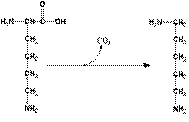
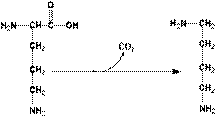
ГНИЕНИЕ БЕЛКОВ В КИШЕЧНИКЕ Около 5% невсосавшихся аминокислот в нижних отделах тонкого и в толстом кишечнике подвергается действию энзимов гнилостной флоры – гниению. В основе гниения лежат реакции декарбоксилирования и дезаминирования с образованием множества продуктов гниения. Кадаверин и путресцин – трупные яды (рис. 2). Обнаруживаются в больших количествах при гастроэнтерите, холере.
Рис. 2. Схема гниения лизина и орнитина
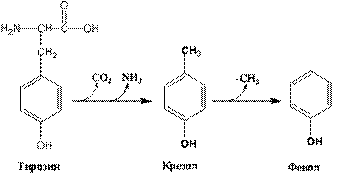
В ходе гниения тирозина и триптофана исходная молекула утрачивает радикал целиком (рис. 3, 4).
Рис. 3. Схема гниения тирозина
Рис. 4. Схема гниения триптофана Продукты гниения аминокислот являются токсичными. Они частично выводятся, а частично всасываются в толстом кишечнике, в последующем обезвреживаются в печени и выводятся с мочой. Для уменьшения количества образующихся продуктов гниения необходимо, чтобы потребление белков соответствовало физиологическому состоянию организма (физическая активность, возраст и т.д.). Кроме того, потребление пищевых волокон (целлюлоза, лигнин, пектин и т.д.) в составе растительных продуктов приводит к подкислению среды в результате их расщепления микрофлорой с образованием кислых продуктов. Кислая среда препятствует процессу гниения. Пищевые волокна улучшают также моторную функцию кишечника, что ускоряет опорожнение толстого кишечника и уменьшает всасывание продуктов гниения.
ОБЩАЯ ХАРАКТЕРИСТИКА КАТАБОЛИЗМА АМИНОКИСЛОТ В процесс катаболизма (распада) вовлекаются только те амии-нокислоты, которые оказались не использованными в ходе реакций синтеза белка или образования других соединений. Катаболизм аминокислот включает два этапа (рис. 5): 1. Дезаминирование, заключающееся в отщеплении аминогруппы в виде аммиака с образованием a-кетокислоты. 2. Катаболизм углеродного скелета аминокислоты, т.е. a-ке-токислоты.
Рис. 5. Схема катаболизма аминокислот
Обязательным этапом катаболизма аминокислот является удаление аминогруппы, т.е. их дезаминирование. Азот аминогруппы выводится у человека в виде мочевины и аммонийных солей. Эти вещества являются конечными продуктами азотистого обмена. Частично аммиак, образующийся при дезаминировании аминокислот, реутилизируется в ходе восстановительного аминирования кетокислот. Углеродные скелеты аминокислот в конечном итоге поступают в ЦТК, где они расщепляются до углекислого газа и воды. Например, при дезаминировании глутамата образуется 2-кетоглутарат, аспартата – оксалоацетат, они являются интермедиатами ЦТК. Катаболизм аланина связан с образованием пирувата, который подвергается окислительному декарбоксилированию с образованием ацетил-КоА, вовлекаемый в ЦТК. Безазотистые углеродные скелеты практически всех протеиногенных аминокислот (за исключением лейцина и лизина) могут служить субстратами глюконеогенеза, т.е. катаболизм некоторой части аминокислот связан с превращением в глюкозу, которая окисляется ради образования энергии. Часть аминокислот, метаболизм которых связан с образованием ацетил-КоА или ацетоацетил-КоА, катаболизируют путем превращения в кетоновые тела или жирные кислоты. К кетогенным аминокислотам относятся фенилаланин, лизин, тирозин, лейцин, изолейцин и триптофан.
ЗАНЯТИЕ 2.
ОБЩАЯ ХАРАКТЕРИСТИКА ПУТЕЙ ОБРАЗОВАНИЯ И ОБЕЗВРЕЖИВАНИЯ АММИАКА
Пути образования аммиака 1) дезаминирование аминокислот 2) распад пуриновых и пиримидиновых оснований 3) гидролиз амидов 4) дезаминирование биогенных аминов Аммиак является высокотоксичным соединением для животных и большинства растений. Особенно к нему чувствительна ЦНС высших животных и человека, на которую он действует как возбуждающий агент, а в повышенных концентрациях как судорожный яд. Образование аммиака в тканях идет непрерывно, но тем не менее он не накапливается в них и его концентрация в крови удерживается на низком уровне (10-20 мкг/100 мл). Аммиак в тканях подвергается связыванию с образованием нетоксичных продуктов, легко выделяемых с мочой. В процессе эволюции наш организм унаследовал 5 основных путей обезвреживания аммиака. 1.Орнитиновый цикл образования мочевины 2.Восстановительное аминирование кетокислот 3.Образование аммонийных солей 4.Синтез креатина (выделяется в виде креатинина с мочой) 5. Образование амидов
ТРАНСАМИНИРОВАНИЕ. Трансаминирование – обратимый перенос аминогруппы с аминокислоты на a-кетокислоту без промежуточного выделения аммиака, в результате чего образуется новая аминокислота и новая кетокислота. Трансаминирование осуществляется пиридоксальзависимыми ферментами – трансаминазами, локализованными как в митохондриях, так и в цитозоле клеток различных органов и тканей. Наиболее активно процесс трансаминирования идет в печени и мышцах. Трансаминированию подвергаются все аминокислоты за исключением пролина, треонина и лизина. В качестве кетокислот, акцептирующих аминогруппу, могут выступать 2-кетоглутарат, оксалоацетат и пируват. Трансаминазы проявляют специфичность по отношению к своей аминокислоте и своей кетокислоте. Ферменты относятся ко второму классу. Наибольшую активность проявляют аланинаминотрансфераза и аспартатаминотрансфераза. Трансаминирование происходит в две стадии. На первой стадии аминокислота взаимодействует с пиридоксальфосфатом в составе фермента, образуя с ним Шиффово основание, отдает ему свою аминогруппу и в результате аминокислота превращается в соответствующую кетокислоту, а пиридоксальфосфат в пиридоксаминофосфат. На второй стадии пиридоксаминофосфат взаимодействует с одной из трех a-кетокислот и передает ей аминогруппу, в результате кетокислота превращается в аминокислоту (2-кетоглутарат в глутамат, оксалоацетат в аспартат, пируват в аланин), пиридоксаминофосфат в пиридоксальфосфат. Процесс является обратимым. Значение трансаминирования: 1.Трансаминирование является первой фазой непрямого дезаминирования. При этом в роли акцептора аминогруппы выступает 2-кетоглутарат, который по ходу реакции превращается в глутамат. Глутамат является единственной аминокислотой, которая может вступать в реакцию окислительного дезаминирования. Таким образом, реакция трансаминирования выполняет функцию своеобразного коллектора, с помощью которого аминогруппы со всех аминокислот собираются на 2-кетоглутарат с образованием глутамата. Поскольку удаление аминогруппы является обязательным условием катаболизма аминокислот эта реакция обеспечивает вовлечение практически всех аминокислот на путь распада. 2.Трансаминирование - один из путей синтеза заменимых аминокислот. 3.Трансаминирование является важнейшим механизмом взаимосвязи обмена различных классов соединений (например, пируват – продукт обмена углеводов в реакции трансаминирования превращается в аланин).
АМИНИРОВАНИЕ КЕТОКИСЛОТ. Основными типами дезаминирования аминокислот являются окислительное и непрямое дезаминирование. Окислительное дезаминирование – процесс отщепления аминогруппы от аминокислоты в виде аммиака с образованием соответствующей кетокислоты. Окислительному дезаминированию подвергается лишь глутамат, поскольку только для этой аминокислоты имеется специфичная высокоактивная НАД(Ф)-зависимая глутаматдегидрогеназа. Этот фермент локализован как в митохондриях, так и в цитозоле клетки. Реакция протекает в две стадии. На первой стадии происходит ферментативное дегидрирование аминокислоты с образованием иминокислоты. Акцептором атомов водорода является НАД(Ф). На второй стадии иминокислота спонтанно, с участием воды, распадается на аммиак и альфа-кетоглутарат. Продукт реакции может включаться в реакции ЦТК либо использоваться в качестве акцептора аминогруппы в реакциях трансаминирования. Реакция имеет значение также для катаболизма таких аминокислот как пролин, глутамин, аргинин и гистидин, так как их метаболизм происходит через стадию образования глутамата. Дезаминирование других аминокислот является непрямым и происходит в два этапа. На первом этапе аминокислоты подвергаются трансаминированию. Причем, в данном случае акцептором аминогруппы служит 2-кетоглутарат. В результате трансаминирования аминокислота лишается аминогруппы, а ее углеродный скелет подвергается катаболизму. В результате присоединения аминогруппы к 2-кетоглутарату образуется глутамат, который вступает во второй этап непрямого дезаминирования, который заключается в его окислительном дезаминировании. Непрямое дезаминирование является наиболее распространенным типом дезаминирования..
ОБМЕН АММИАКА Основным источником аммиака является дезаминирование аминокислот. Аммиак образуется также в при распаде биогенных аминов и нуклеотидов, гидролизе глутамина и аспарагина. Существенная часть аммиака образуется в кишечнике в результате деятельности микрофлоры и поступает в кровь воротной вены. Образование аммиака происходит во всех тканях, однако его концентрация поддерживается на довольно низком уровне (25–40 мкмоль/л), так как он довольно быстро обезвреживается. Существование механизмов обезвреживания аммиака связано с его токсичностью. Механизмы токсичности аммиака 1.Аммиак сдвигает реакцию восстановительного аминирования кетокислот в сторону образования глутамата 2-кетоглутарат + НАДН2 + NH3 = глутамат + НАД Снижение концентрации a-кетоглутарата вызывает угнетение обмена аминокислот, так как снижается количество акцептора в реакциях трансаминирования, а также гипоэнергетическое состояние в связи со снижением скорости ЦТК. 2.Аммиак усиливает синтез глутамина из глутамата в нервной ткани накопление глутамина в нервных клетках приводит к повышению осмотического давления и может вызвать отек мозга. Снижение концентрации глутамата нарушает образование ГАМК – основного тормозного медиатора Существует несколько способов обезвреживания и выведения аммиака в разных тканях. Основной реакцией обезвреживания аммиака почти во всех тканях является синтез глутамина под действием глутаминсинтазы. глутамат + NH3 + АТФ = глутамин + АДФ + н. Фосфат Глутамин является транспортной формой аммиака, так как представляет собой аминокислоту, способную легко проникать через клеточные мембраны. В наибольших количествах глутамин образуется в мышцах и мозге. Образовавшийся в тканях глутамин транспортируется в печень и почки, где он подвергается действию глутаминазы и расщепляется с образованием глутамата и аммиака. Аммиак в печени вовлекается в реакции орнитинового цикла, в котором образуется мочевина, в почках он используется для образования аммонийных солей. Глутаминаза почек активируется при ацидозе, полученный аммиак используется для образования аммонийных солей, что позволяет нейтрализовать кислые продукты обмена. Экскреция солей аммония составляет 0,5 г/сут. Эта реакция защищает также организм от потери натрия и калия, которые могли бы использоваться в качестве катионов при образовании солей с анионами кислот. Еще одной реакцией обезвреживания аммиака в мозге и некоторых других тканях является восстановительное аминирование 2-кетоглктарата под действием глутаматдегидрогеназы, катализирующей обратимую реакцию. Однако этот путь обезвреживания аммиака используется слабо хотя и представляется выгодным для клеток, так как способствует обезвреживанию сразу 2 молекул аммиака. 2-кетоглутарат +NH3 = глутамат + NH3 = глутамин
ОРНИТИНОВЫЙ ЦИКЛ МОЧЕВИНООБРАЗОВАНИЯ
Орнитиновый цикл мочевинообразования (цикл Кребса-Ген-зелейта) является главным механизмом обезвреживания аммиака в организме. Осуществляется только в клетках печени. Основная часть аммиака для орнитинового цикла поставляется из кишечника, а также при дезаминировании аминокислот в печени и гидролиза глутамина. В ходе этого цикла образуется мочевина – основной конечный продукт азотистого обмена. На долю мочевины приходится 80–85% всего азота, выводимого из организма. Мочевина – нейтральное, хорошо растворимое в воде соединение, легко удаляется из организма с мочой. В среднем за сутки с мочой взрослого человека выводится около 30 г мочевины. Орнитиновый цикл – процесс ферментативный, энергозависимый. На образование одной молекулы мочевины расходуется 3 АТФ. Начальные этапы цикла осуществляются в митохондриях, промежуточные и заключительные – в цитозоле клетки. Орнитиновый цикл начинается с взаимодействия свободного аммиака с углекислым газом, в результате которого образуется карбомоилфосфат. Реакция катализируется карбомоилфосфаттрансферазой и требует присутствия двух молекул АТФ. Одна из них используется в качестве донора фосфатной группы, включаемой в состав карбомоилфосфата. Карбомоилфосфат далее конденсируется с орнитином с образованием цитруллина. Орнитин в данной реакции выступает в качестве своеобразного катализатора, так как от его количества зависит скорость всего цикла. Цитруллин поступает в цитоплазму, где взаимодействует с аспартатом, в результате чего образуется аргининосукцинат. Аспартат приносит в состав мочевины еще одну аминогруппу. Аргининосукцинат в следующей реакции цикла расщепляется с образованием фумарата и аргинина. Аргинин под действием аргиназы расщепляется с образованием мочевины и орнитина. Мочевина выводится из организма с мочой, а орнитин вступает в новый цикл мочевинообразования. Фумарат, образовавшийся в предыдущей реакции, в реакциях ЦТК превращается до оксалоацетата, который в реакции трансаминирования акцептирует аминогруппу аминокислоты и превращается в аспартат и участвует в реакции с цитруллином. Источником азота одной аминогруппы мочевины является свободный аммиак, источником второй – аспартат. Между орнитиновым циклом мочевинообразования и циклом трикарбоновых кислот существует тесная функциональная зависимость. ЦТК поставляет в орнитиновый цикл СО2 и АТФ. При участии ферментов ЦТК происходит превращение фумарата в аспартат, необходимый для образования аргининосукцината. Последовательность реакций превращения фумарата в аспартат, который включается в орнитиновый цикл, выполняет роль своеобразного привода, обеспечивающего функционирование двух циклических процессов: ЦТК и орнитинового цикла.
ЗАНЯТИЕ № 3 СПЕЦИФИЧЕСКИЕ ПУТИ ОБМЕНА АМИНОКИСЛОТ. ОБМЕН НУКЛЕОТИДОВ. ВЗАИМОСВЯЗЬ ОБМЕНОВ ВЕЩЕСТВ
БИОГЕННЫЕ АМИНЫ: ОБРАЗОВАНИЕ И ИНАКТИВАЦИЯ
Биогенные амины являются продуктами декарбоксилирования ряда аминокислот (тирозина, триптофана, гистидина, глутамата) или их гидроксипроизводных (дигидроксифенилаланина – ДОФА, 5-гидрокситриптофана). К биогенным аминам относятся катехоламины (дофамин, норадреналин, адреналин), гистамин, серотонин, триптамин, тирамин, ГАМК (гамма-аминомасляная кислота). Образование биогенных аминов происходит в специализированных клетках (например, катехоламины образуются в клетках мозгового слоя надпочечников, гистамин – в тучных клетках и т.д.) под влиянием специфических тканевых В6-витаминозависимых декарбоксилаз. Синтезированные биогенные амины накапливаются в клетке в комплексе с белками или нуклеотидами. Под влиянием внешнего стимула происходит высвобождение биогенных аминов из связанных форм и активный выход из клетки путем экзоцитоза. Биогенные амины, обладая высокой физиологической активностью, могут выполнять следующие функции: а) нейромедиаторов: дофамин, норадреналин, гистамин, серотонин, триптамин, ГАМК; б) истинных гормонов: адреналин, норадреналин; в) тканевых гормонов: гистамин, серотонин. Накопление биогенных аминов могло бы отрицательно сказаться на течении физиологических процессов и привести к серьезным нарушениям регуляции обменных процессов в отдельных тканях и организме в целом. Однако органы и ткани организма обладают специальными механизмами инактивации биогенных аминов. Основным механизмом инактивации биогенных аминов является их окислительное дезаминирование под влиянием фермента – моноаминооксидазы. Образующиеся в результате окислительного дезаминирования альдегиды окисляются до соответствующих кислот. Моноаминооксидаза является ФАД-содержащим ферментом и локализуется преимущественно в митохондриях. Некоторые ингибиторы моноаминооксидазы используются в медицине для лечения гипертонической болезни, депрессии, шизофрении. Гистамин образуется при декарбоксилировании гистидина, является нейромедиатором. Вызывает расширение кровеносных сосудов, понижает артериальное давление, способствует притоку лейкоцитов к местам воспаления (защитная реакция организма против инфекции). Участвует в процессах сенсибилизации организма (поэтому аллергические состояния лечат противогистаминными препаратами). Способствует отделению соляной кислоты в желудочном соке. ГАМК образуется при декарбоксилировании глутамата. Реакция происходит под действием глутаматдекарбоксилазы, локализованной в сером веществе мозга. Является нейромедиатором, оказывает выраженное тормозное влияние на центральную нервную систему. Используется как компонент лекарственных средств, применяемых при эпилепсии, а также при заболеваниях с выраженным моторным компонентом. Серотонин образуется при декарбоксилировании 5-гидрокситриптофана, является мощным сосудосуживающим агентом и стимулятором сокращения гладких мышц. Серотонин известен как важный нейромедиатор, участвующий в восприятии болевых раздражений (и в блокировке болевой чувствительности в экстремальных ситуациях), координации моторной активности, эмоциональном поведении, поддержании ритма сна и бодрствования (наряду с мелатонином, производным серотонина), терморегуляции, а также во многих других процессах. Серотонин регулирует кишечную перистальтику, вызывает сокращение мускулатуры матки, бронхов и других гладкомышечных органов у животных и человека. Катехоламины образуются в мозговом слое надпочечников, а также в клетках мозга. Предшественником в синтезе является тирозин, который в первой реакции подвергается гидроксилированию с образованием ДОФА – дигидроксифенилаланина. В дальнейшем ДОФА декарбоксилируется и образуется дофамин. b-гидроксилирование дофамина приводит к образованию норадреналина, который в реакции трансметилирования (донором метильной группы выступает S-аденозилметионин) превращается в адреналин. В клетках мозга отсутствует фермент трансметилирования, поэтому образуется только норадреналин. Адреналин и норадреналин являются истинными гормонами. Они обладают сосудосуживающим эффектом, расширяют просвет бронхов, увеличивают частоту сердечных сокращений, способствуют мобилизации жира и гликогена из депо, усиливают гликогенолиз в мышцах.
ТРАНСМЕТИЛИРОВАНИЕ Трансметилирование – это процесс переноса метильной группы (СН3) от метионина, который содержит лабильную метильную группу. Метионин участвует в этом процессе в своей активной форме – в виде S-аденозилметионина, образующегося в реакции с АТФ. S-аденозилметионин под влиянием метилтрансферазы передает свою СН3-группу на вещество, подлежащее метилированию, т.е. акцептору. Сам он при этом превращается в S-аденозилгомоцистеин, а акцептор становится метилированным. Далее S-аденозилгомоцистеин расщепляется на аденозин и гомоцистеин. Наиболее энергично процесс трансметилирования протекает в митохондриях клеток печени. Биороль трансметилирования
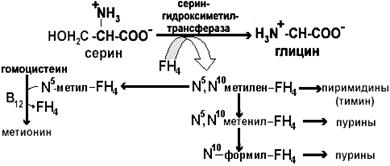
Трансметилирование используется в процессе синтеза адреналина из норадреналина, образования креатина, фосфатидилхолина из фосфатидилэтаноламина, холина из этаноламина, посттрансляционной модификации полипептидных цепей белка, процессинга рибонуклеиновых кислот (расстановка метильных меток в РНК обеспечивает их защиту от собственных эндо- и экзонуклеаз). Трансметилирование является важнейшим механизмом обезвреживания ксенобиотиков путем конъюгации аминов, фенолов и тиоловых соединений с образованием метилпроизводных. Образовавшийся в реакции трансметилирования гомоцистеин может повреждать эндотелий сосудов, в связи с чем повышенная концентрация рассматривается как этиологический фактор развития атеросклероза. Путями утилизации гомоцистеина в организме являются Вс- и В12-зависимое метилирование с образованием метионина и В6-зависимое образование цистеина. Донором метильной группы в первом случае выступает метил-ТГФК. Для успешного протекания реакции требуется присутствие кобаламина, который является коферментом метилтрансферазы. Метильная группа вначале переносится на кобаламин, а уже с метилкобаламина на гомоцистеин. Таким образом, в регенерации метионина задействованы два витамина. Функция фолиевой кислоты шире, чем участие только в реакциях трансметилирования. Она обеспечивает обмен и других одноуглеродных фрагментов – формильных, формимино, метинильных, оксиметильных и метиленовых. Основным источником одноуглеродных фрагментов является серин, который, взаимодействуя с активной формой фолиевой кислоты – тетрагидрофолиевой кислотой (FН4), передает ей метиленовую группу (рис. 1). Метилен в составе FH4 либо восстанавливается до метил-FH4, который затем переносится на гомоцистеин с образованием метионина, либо используется в реакциях синтеза тимина. Другой возможный путь – превращение метилен-FH4 в метинил-FH4 или в формил-FH4, которые используются в реакциях синтеза пуриновых нуклеотидов.
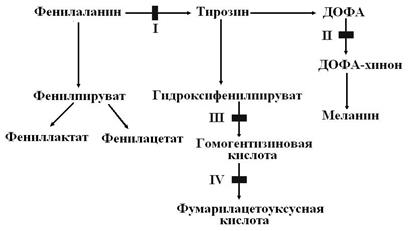
Рис.1 Роль фолиевой кислоты в обмене одноуглеродных фрагментов НАСЛЕДСТВЕННЫЕ НАРУШЕНИЯ ОБМЕНА АМИНОКИСЛОТ Наиболее часто встречаются четыре вида молекулярных болезней, связанных с обменом фенилаланина и тирозина. Их причиной являются метаболические блоки на разных этапах обмена этих аминокислот (рис. 2).
Рис. 2 Метаболические блоки ТКАНЕВЫЕ НУКЛЕАЗЫ Нуклеиновые кислоты поступают с пищей в составе нуклеопротеинов. Их расщепление на составные части – нуклеиновые кислоты и белковый компонент – осуществляется в желудке под действием соляной кислоты и пепсина. Белковая часть нуклеопротеинов расщепляется протеиназами пищеварительного тракта. Нуклеиновые кислоты ДНК и РНК расщепляются путем гидролиза межнуклеотидных 3’, 5’-фосфорнодиэфирных связей в тонком кишечнике под действием ДНК-азы (дезоксирибонуклеазы) и РНК-азы (рибонуклеазы) поджелудочного сока соответственно с образованием олигонуклеотидов. Последние под действием фосфодиэстераз поджелудочной железы расщепляются до мононуклеотидов. Нуклеотиды лишаются фосфатного остатка под действием нуклеотидаз и фосфатаз и превращаются в нуклеозиды, которые либо всасываются слизистой кишечника, либо расщепляются нуклеозидфосфорилазами кишечника до оснований и дезокси- или рибозо-1-фосфата. Пищевые пурины и пиримидины не являются незаменимыми факторами пищи, так как практически не используются для синтеза нуклеиновых кислот. Это связано с высокой активностью ферментов, осуществляющих деградацию оснований до конечных продуктов обмена еще в энтероцитах. Что касается тканевых нуклеаз, то они осуществляют расщепление нуклеиновых кислот аналогично нуклеазам поджелудочного сока. Существуют нуклеазы двух типов: сайт-неспецифические нуклеазы (ДНК-азы и РНК-азы) и сайт-специфические нуклеазы – ре-стриктазы. Отличаются они тем, что сайт-специфические рестриктазы гидролизуют фосфорнодиэфирные связи в строго определенных последовательностях из 6–8-ми нуклеотидов. Для каждой рестриктазы существует своя последовательность нуклеотидов. Распад чужеродной (в основном фаговой) ДНК под действием рестриктаз также происходит в строго определенных зонах молекулы, имеющих структуру палиндромов. Поскольку рестриктазы действуют на строго определенные участки молекулы, они используются для расшифровки последовательности нуклеотидных остатков в ДНК фагов и вирусов. Более того, это уникальное свойство рестриктаз находит все боль
|
|||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-12; просмотров: 630; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.151 (0.019 с.) |