Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
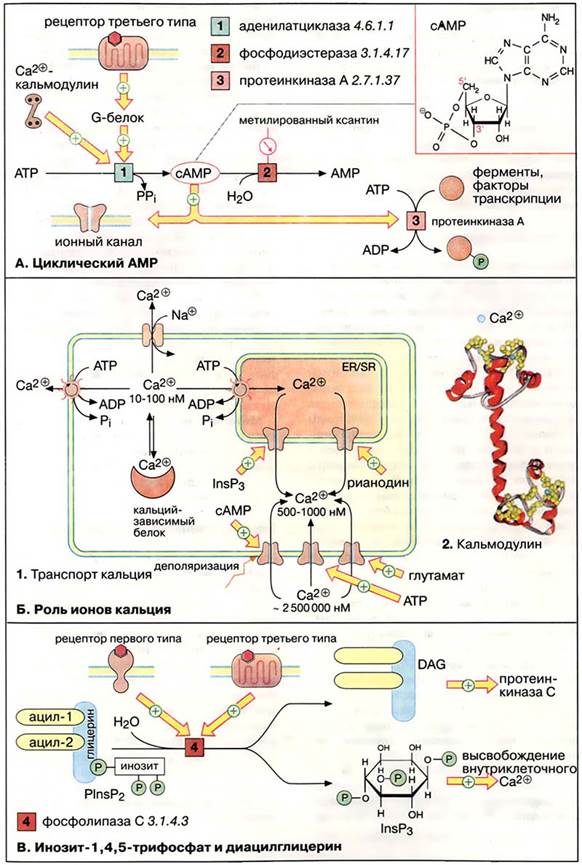
Механизмы передачи информации от внешних сигналов на внутриклеточные процессы. Роль ионов кальция в механизмах трансформации внешних сигналов. Калмодулин.Содержание книги
Поиск на нашем сайте
Уровень ионов кальция. Концентрация ионов Са2+ в цитоплазме нестимулированной клетки очень низка (10-100 нМ). Низкий уровень поддерживается кальциевыми АТФ-азами (кальциевыми насосами) и натрий-кальциевыми обменниками. Резкое повышение концентрации ионов Са2+ в цитоплазме (до 500-1000 нМ) происходит в результате открывания кальциевых каналов плазматической мембраны или внутриклеточных кальциевых депо (гладкого и шероховатого эндоплазматического ретикулума). Открывание каналов может быть вызвано деполяризацией мембран или действием сигнальных веществ, нейромедиаторов (глутамат и АТФ, см. Медиаторы нервной системы), вторичных мессенджеров (ИФ3 и цАМФ), а также вещества растительного происхождения рианодина. В цитоплазме и клеточных органеллах имеется множество белков способных связывать Са2+, некоторые из них выполняют роль буфера. При высокой концентрации в цитоплазме ионы Са2+ оказывает на клетку цитотоксическое действие. Поэтому уровень кальция в отдельной клетке испытывает кратковременные всплески, увеличиваясь в 5-10 раз, а стимуляция клетки увеличивает лишь частоту этих флуктуаций. Действие кальция опосредовано специальными Са2+-связывающими белками («кальциевыми сенсорами»), к которым принадлежат аннексии, кальмодулин и тропонин (см. Регуляция сокращения мышечных волокон). Кальмодулин — сравнительно небольшой белок (17 кДа) — присутствует во всех животных клетках. При связывании четырёх ионов Са2+ (на схеме голубые кружочки) кальмодулин переходит в активную форму, способную взаимодействовать с многочисленными белками. За счёт активации кальмодулина ионы Са2+ оказывают влияние на активность ферментов, ионных насосов и компонентов цитоскелета.
--- 38. Роль ограниченного протеолиза в механизмах регуляции процессов жизнедеятельности. Свертывание крови. Факторы и механизмы свертывания. Значение ионов кальция и витамина К в процессах свертывания крови. Ограниченный (частичный) протеолиз проферментов подразумевает, что синтез некоторых ферментов осуществляется в виде более крупного предшественника и при поступлении в нужное место этот фермент активируется через отщепление от него одного или нескольких пептидных фрагментов. Подобный механизм защищает внутриклеточные структуры от повреждений.
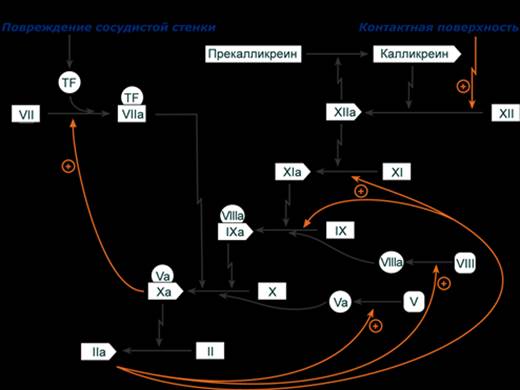
Примером служит активация протеолитических ферментов желудочно-кишечного тракта (трипсиноген, пепсиноген, прокарбоксипептидазы), факторов свертывающей системы крови, лизосомальных ферментов (катепсины). Секреция ряда ферментов за пределы клетки в неактивном состоянии позволяет предохранить клетки от повреждения (пищеварительные ферменты) или сохранить белок в плазме крови до наступления определенного момента (факторы свертывания крови, белки системы комплемента, калликреин-кининовой и ренин-ангиотензиновой систем). При повреждении крупных кровеносных сосудов тромбоцитарная пробка не способна остановить кровотечение, она относительно рыхлая и непрочная. Только коагуляционный гемостаз способен остановить кровотечение из такого сосуда. В классической теории коагуляции выделяют 2 пути активации факторов свертывания: 1. Активация тканевым фактором. Так как тканевой фактор не относится к плазменным факторам и контактирует с кровью только при повреждении сосуда, то активация с его участием обозначается как внешний путь свертывания. 2. Контактная активация – активация фактора ХII при взаимодействии с отрицательно заряженной поверхностью (in vitro) или при воспалении (in vivo). Поскольку фактор XII в норме присутствует в плазме, активация с его участием обозначается как внутренний путь свертывания. Внешний и внутренний пути сходятся на Х факторе, активная форма которого Xa, вместе с фактором Va и Са2+ формирует ферментативный комплекс Xa-Va-Ca2+, иначе называемый протромбиназа, которая превращает протромбин в тромбин. Образовавшийся тромбин превращает фибриноген в фибрин-мономеры. Последние соединяются, образуя полимеры фибрина. Реакции с участием VII, IX, X, XI, XII факторов происходят на фосфолипидной поверхности (тромбопластине). Внешний путь свертывания После повреждения сосуда тканевой фактор (TF), находящийся на клетках, связывает и активирует фактор VII. Образованный комплекс напрямую активирует фактор Х. Далее ф.Ха при участии кофактора Va в присутствии ионов Са2+ формирует комплекс Xa-Va-Ca2+ – протромбиназу, превращающую протромбин в тромбин. Активность внешнего пути поддерживается за счет механизма положительной обратной связи: - образующийся тромбин активирует V фактор, - фактор Xa в присутствии ионов Cа2+ активирует фактор VII.
Внутренний путь свертывания Внутренний путь свертывания развертывается на фосфолипидной поверхности тромбоцитов или иных клеток, где в первую очередь происходит сборка комплекса, состоящего из факторов XII, XI, прекалликреина и высокомолекулярного кининогена (ВМК). 1. Активация фактора XII. Связывание фактора XII с тромбопластином (тканевым фактором) изменяет его конформацию, и он приобретает небольшую активность. Это позволяет фактору XIIа начать превращение прекалликреина в калликреин. Затем, в результате действия калликреина накапливается фактор XIIa, и активация калликреина усиливается. Т.о. фактор XIIа и калликреин взаимно активируют друг друга. Также фактор XII может активироваться фактором VIIa. 2. Активация фактора XI. Фактор XIIa активирует фактор XI. 3. Активация фактора IX. Фактор XIa в присутствии ионов Са2+ локализует на мембране и активирует фактор IX. Фактор IX может также активироваться фактором VIIa. Далее фактор IXa связывается со своим кофактором VIIIа и образует комплекс IXa-VIIIa-Са2+, называемый теназа или теназный комплекс (англ. ten - десять). 4. Активация фактора X. Теназа (комплекс IXa-VIIIa-Са2+) активирует фактор X. Активированный фактор Ха при помощи своего кофактора Va в присутствии ионов Са2+ на фосфолипидной мембране формирует комплекс Xa-Va-Ca2+ – протромбиназу. ---
5. Активация фактора II (тромбина). Протромбиназа атакует протромбин и последовательно расщепляет две связи в его молекуле, отделяя N-концевой фрагмент, с формированием активного тромбина.
6. Тромбин
превращает фибриноген в фибрин-мономер, по мере своего образования через обратные положительные связи активирует факторы V, VIII, XI, что поддерживает активность ферментативного каскада. ---
|
||||
|
|
Последнее изменение этой страницы: 2021-05-12; просмотров: 344; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.20 (0.007 с.) |