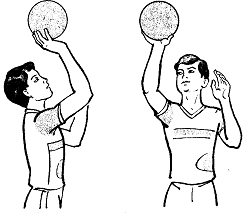
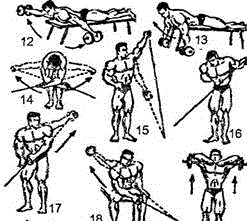
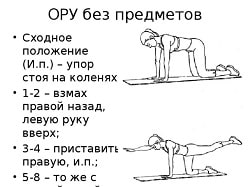
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Принципы кодирования номером каналаСодержание книги
Поиск на нашем сайте Понятие анализатора, введенное И.П. Павловым, в процессе изучения нейронных механизмов сенсорных систем пополнилось новым содержанием. Оказалось, что сигналы, выделенные рецепторами, подвергаются анализу в нейронных сетях так, что отдельные нейроны выделяют дробные признаки. Тот факт, что разные параметры сигнала представлены возбуждением различных нейронов, означает, что анализ осуществляется посредством кодирования сигнала номером канала. Кодирование сигнала рисунком разряда одного нейрона и кодирование ансамблем нейронов не имеют надежных экспериментальных подтверждений. Принцип кодирования номером канала состоит в том, что: 1) определенному сигналу в нервной системе ставится в соответствие определенный максимально возбужденный нейрон-детектор, 2) определенной реакции соответствует максимальное возбуждение реализующего ее командного нейрона. Таким образом при изменении стимула или при смене реакций максимум возбуждения перемещается с одного детектора или командного нейрона на другой. В эволюции кодирование сигнала номером канала ведет свое начало от кожной чувствительности, где изменение положения стимула приводит к перемещению максимума возбуждения с одного участка рецептивной поверхности на другой. Дальнейшее усложнение этой схемы представлено в слуховом анализаторе, где звук не может непосредственно перемещаться по рецептивной поверхности и кодирование номером канала осуществляется смещением места, в котором достигается максимум деформации волосковых клеток внутреннего уха при изменении частоты и интенсивности звука. В том случае, когда стимул не может вызывать даже опосредованного смещения максимума возбуждения по рецептивной поверхности, кодирование номером канала реализуется за счет такой системы связей между рецепторами и афферентными нейронами, при которой определенной комбинации возбуждений рецепторов соответствует единственный максимум возбуждения на наборе афферентных нейронов. При изменении стимула меняется комбинация возбуждений рецепторов и максимум возбуждения перемещается с одного афферентного нейрона на другой. Примером может служить цветовое зрение, где изменение комбинаций возбуждений трех типов колбочек приводит к перемещению максимума возбуждения с одного цветоселективного афферентного нейрона на другой. 7. Механизмы научения Исторически сложились два конфликтующих между собой подхода к проблеме обучения. Сетевой подход восходит к нейронной доктрине. В ней подчеркивается роль нейрона как функциональной единицы нервной системы, а синапсам отводится решающая роль в хранении информации. В своей ранней версии этот подход основывался на нейробиотаксисе, который использовался для того, чтобы объяснить некоторые факты нейроэмбриологии и сравнительной анатомии. В соответствии с концепцией нейробиотаксиса, рост новых синапсов стимулируется продолжительной электрической активностью двух нервных клеток, что становится причиной прорастания терминалей аксона одной клетки к дендритам другой, тем самым формируя новый синапс. «Полевой» подход восходит к гештальт-теории. Предполагается, что информация хранится в биоэлектрических полях, создаваемых аггрегированной активностью многих нейронов. Гештальт-теория подчеркивала, что восприятие зависит от конфигурации в целом, а не от суммы каких-то отдельных частей. Аналогичным же образом в «полевой» гипотезе подчеркивался глобальный аспект нейронной активности, а индивидуальный вклад нейронов и специфика нейронных соединений игнорировалась совсем. Начиная с 20-х гг. XX в., наиболее ярким выразителем идей «полевого» подхода был К. Лешли. В результате своих исследований, связанных с экстирпацией участков коры головного мозга и изучением влияния этих операций на обучение в лабиринте, он сделал вывод о том, что поведенческие нарушения зависят от количества экстирпированного мозгового вещества, а не от локализации повреждения. К. Лешли считал, что будущее развитие клеточной нейрофизиологии мало поможет изучению механизмов памяти и обучения. Сейчас, критически оценивая эти взгляды, не стоит забывать о том, что нейрофизиология во времена К. Лешли еще толь- 60 Глава 1. Нейрофизиология. Клеточные основы обучения 7. Механизмы научения 61
Для ответа на вопрос о том, обладает ли система памятью и каковы ее характеристики, в простейшем-случае можно использовать многократное повторение внешнего сигнала с интервалами, превосходящими время переходного процесса. Если в результате повторений сигнала система оказывается способной воспроизвести запись, то она обладает механизмом записи, сохранения, считывания и воспроизведения, памятью в собственном смысле слова. Общая характеристика памяти включает длительность процесса фиксации, прочность удержания, продолжительность и объем сохранения, полноту, точность и скорость считывания и особенности воспроизведения. Следовые процессы При изучении реакции на уровне отдельного нейрона следует иметь в виду, что термин «отдельный нейрон» относится к условиям микроэлектродного отведения. Реально работа изучаемого нейрона определяется совокупностью нейронов, образующих нейронную сеть. Поскольку разные нейроны занимают разное место в нейронной сети, их реакции будут существенно отличаться даже при полной идентичности их внутренних свойств. В реакциях изучаемого нейрона отражается сложное взаимодействие свойств нейронной сети и специфических особенностей данного элемента. При классификации следовых процессов необходимо рассматривать их зависимость от отсчета времени и возможности воспроизведения по сигналам, поступающим от других рецепторов. Первый критерий позволяет выделить следовые эффекты, связанные и не связанные с отсчетом времени после действия раздражителя. Первая группа включает: а) следовой эффект в виде переходного процесса, время которого определяется возвращением спайковой активности к исходному уровню. В ряде случаев спонтанная ритмика у нейрона отсутст- вует, поэтому время возвращения к исходному состоянию обнаружить не удается. В этом случае использование парных раздражений позволяет косвенно исследовать фазы переходного процесса; б) с/гедовые эффекты в виде изменения свойств переходного процесса: длительности, величины, латентного периода, конфигурации. Наконец, важным показателем следовых эффектов может быть вариативность реакции и, в частности, ее латентного периода, а также отношение сигнал-шум. Во вторую группу следов входят эффекты, связанные с отметкой момента действия стимула: а) угасание реакций, избирательное в отношении момента раздражения; б) угасание реакций, специфическое в отношении интервала между раздражениями усиления или появление реакции. В данном случае речь идет о нейронном уровне условного рефлекса на время: тормозного или возбудительного. Используя получение информации от других анализаторов как критерий разделения следовых процессов, следует рассмотреть динамические свойства разных типов конвергенции сигналов на одном нейроне. Развитие методов нейрофизиологического анализа работы нервных клеток позволило исследователям подойти к интерпретации явлений памяти. Обычно данные о процессах памяти получают на основе результатов воспроизведения. Однако воспроизведение само зависит от условий восприятия, условий фиксации, избирательности считывания, особенностей эфферентной системы, участвующей в воспроизведении сигнала. Энграммы памяти могут храниться в течение всей жизни, а доступ к ним утрачивается. Опыты, проведенные У. Пенфилдом (1969), показали, что при раздражении электрическим током зон «перекрытия» анализаторов на границе височной, затылочной и теменной областей коры больших полушарий больной, находящийся под местной анестезией, вновь переживает целые сцены из своего прошлого. Эти переживания либо носят характер сновидений, либо приобретают яркость галлюцинаций. Больной слышит мелодии, голоса, узнает говорящих, видит входящих в помещение людей. Проверка показала, что действительно воспроизводятся картины пережитых событий, которые в обычных условиях не вспоминаются. Анализ случаев феноменальной памяти, проведенный А.Р. Лурия, показывает возможность безошибочного воспроизведения сложного бессвязного текста спустя пятнадцать лет после его первоначального заучивания, что также свидетельствует в пользу устойчивости следов памяти. Следовательно, можно говорить о существовании механизма памяти в виде непрерывной записи событий с одновременной отметкой времени каждого переживания. Однако лишь часть этих «записей» поддается в обычных условиях воспроизведению, которое ограничено воз- 62 Глава I Нейрофизиология Клеточные основы обучения 7 Механизмы научения 63
Привыкание Одной из важнейших реакций животных и человека является реакция на новизну стимула — ориентировочный рефлекс, или рефлекс «Что такое?», как назвал его И.П. Павлов. При повторении стимула развивается привыкание. Оно является негативным научением, одной из простейших форм приобретения индивидуального опыта, которое состоит в постепенном избирательном подавлении и полном исчезновении реакции при повторении раздражителя с интервалами, исключающими периферическую адаптацию и эффект утомления. По целому ряду признаков «негативное научение» совпадает с внутренним торможением. Исследование угашения разных компонентов ориентировочного рефлекса показало, что избирательность процесса угашения опосредована следом повторяющегося раздражителя — «нервной моделью стимула». Преимуществом «негативного научения» при исследовании следовых процессов на уровне поведения и отдельного нейрона по сравнению с другими видами условных рефлексов является простота экспериментальной ситуации, сводящейся к многократному нанесению раздражителя. Термин «привыкание» первоначально использовался в поведенческих опытах как на простых, так и на сложных организмах с целью описания снижения ответа на монотонно повторяющийся стимул. Привыкание отличается от утомления тем, что после предъявления экстрастимула — любого стимула, отличающегося от ранее использованного по одному из параметров — наблюдается восстановление ответа (растормаживание). Очевидно, что декремент ответа является свойством многих видов поведения и нейронных объединений. Даже простейшие нейронные системы демонстрируют такой декремент, параметрически сходный с привыканием, которое демонстрируется на уровне интактного организма. Основные параметры привыкания следующие: 1) при повторении стимула наблюдается снижение амплитуды вы 2) чем чаще предъявление стимула, тем быстрее и отчетливее дек 3) на более слабый стимул декремент более отчетливый, на силь 4) восстановление может быть продолжено при помощи повтор 5) привыкание ответа на данный стимул может генерализоваться и 6) при прекращении стимуляции происходит спонтанное восста 7) привыкание происходит быстрее при повторных предъявлениях
8) предъявление сильного экстрастимула вызывает восстановле 9) при повторных использованиях эффективность экстрастимула Ценность этих характеристик привыкания в том, что они применимы и для идентификации явлений снижения ответов у простых нервных систем. Привыкание является наиболее подробно изученной формой стимул-зависимого обучения. Его называют также негативным научением, поскольку главная его задача — научиться не реагировать на опре- 64 Глаиа I Нейрофизиология. Клеточные основы обучения 7 Механизмы научения 65
|
||
|
|
Последнее изменение этой страницы: 2021-01-09; просмотров: 374; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.115 (0.013 с.) |



 ко начиналась. Ответом на «антиконнекционизм» К. Лешли стали опыты в области генетической биологии. Р. Сперри (1940) начал изучать специфичность развития связей во время регенерации нерва и показал, что у низших позвоночных такое сложное поведение, как зрительное восприятие и двигательная координация, самым лучшим образом может быть объяснено с позиций высокоспецифических связей. С тех пор в нейронауке накоплен колоссальный фактический материал, который не обеспечивает безусловной эмпирической опоры ни одной из этих концепций. На самом деле, все новые и новые открытия в области нейрогенетики, нейрохимии и нейрофизиологии еще только позволяют сформулировать вопросы, решение которых станет принципиальным для понимания механизмов обучения и памяти.
ко начиналась. Ответом на «антиконнекционизм» К. Лешли стали опыты в области генетической биологии. Р. Сперри (1940) начал изучать специфичность развития связей во время регенерации нерва и показал, что у низших позвоночных такое сложное поведение, как зрительное восприятие и двигательная координация, самым лучшим образом может быть объяснено с позиций высокоспецифических связей. С тех пор в нейронауке накоплен колоссальный фактический материал, который не обеспечивает безусловной эмпирической опоры ни одной из этих концепций. На самом деле, все новые и новые открытия в области нейрогенетики, нейрохимии и нейрофизиологии еще только позволяют сформулировать вопросы, решение которых станет принципиальным для понимания механизмов обучения и памяти.

 можностями считывания. Используя метод суммирования вызванных потенциалов, отводимых с помощью вживленных в лобные отделы коры больших полушарий человека электродов, Г Уолтер (1965) обнаружил «волну ожидания» в форме негативного колебания, появляющегося всякий раз перед тем, как испытуемому предстояло совершить действие или принять решение. Электроэнцефалограммы (ЭЭГ) позволяют приблизиться к внутренним процессам и получить новые данные о природе нервного следа. Рассмотренные выше эффекты репродукции следа позволят осуществить «наложение» реакции на внешний сигнал и получить данные о «конфигурации» следа памяти. Однако такие исследования памяти ограничены возможностями механизмов считывания и воспроизведения. Исключить или снизить их участие в механизме считывания и воспроизведения при изучении памяти можно при помощи анализа узнавания, основанного на сличении внешних сигналов со следами раздражений, хранимых в памяти. Обычно при изучении узнавания испытуемые выбирают из серии объектов те, которые были им предъявлены ранее. В этом случае механизм воспроизведения следа исключается и эффективность памяти измеряется количеством правильных идентификаций. Однако описанный прием не позволяет исследовать «конфигурацию» следа памяти. Для того чтобы метод узнавания стал пригодным для изучения конфигурации следа памяти, порядок исследования должен быть несколько изменен. Сначала предъявляется стимул-эталон, с которым и сравниваются поступающие позже сигналы. Привыкание и есть одна из форм обучения, результатом которой является формирование энграммы стимула-эталона. Рассогласование действующего стимула с созданной энграм-мой становится причиной развития ориентировочной реакции.
можностями считывания. Используя метод суммирования вызванных потенциалов, отводимых с помощью вживленных в лобные отделы коры больших полушарий человека электродов, Г Уолтер (1965) обнаружил «волну ожидания» в форме негативного колебания, появляющегося всякий раз перед тем, как испытуемому предстояло совершить действие или принять решение. Электроэнцефалограммы (ЭЭГ) позволяют приблизиться к внутренним процессам и получить новые данные о природе нервного следа. Рассмотренные выше эффекты репродукции следа позволят осуществить «наложение» реакции на внешний сигнал и получить данные о «конфигурации» следа памяти. Однако такие исследования памяти ограничены возможностями механизмов считывания и воспроизведения. Исключить или снизить их участие в механизме считывания и воспроизведения при изучении памяти можно при помощи анализа узнавания, основанного на сличении внешних сигналов со следами раздражений, хранимых в памяти. Обычно при изучении узнавания испытуемые выбирают из серии объектов те, которые были им предъявлены ранее. В этом случае механизм воспроизведения следа исключается и эффективность памяти измеряется количеством правильных идентификаций. Однако описанный прием не позволяет исследовать «конфигурацию» следа памяти. Для того чтобы метод узнавания стал пригодным для изучения конфигурации следа памяти, порядок исследования должен быть несколько изменен. Сначала предъявляется стимул-эталон, с которым и сравниваются поступающие позже сигналы. Привыкание и есть одна из форм обучения, результатом которой является формирование энграммы стимула-эталона. Рассогласование действующего стимула с созданной энграм-мой становится причиной развития ориентировочной реакции.

 деленный стимул. Привыкание в той или иной степени присутствует в разных рефлексах. Степень его выраженности зависит от специфичности стимула, силы раздражения и состояния организма. В самом общем виде привыкание — это постепенное снижение реакции в ходе повторений стимула. Отличие привыкания от утомления и истощения в том, что реакцию можно восстановить простым изменением стимула. Наиболее отчетливо привыкание выражено в системе ориентировочного рефлекса.
деленный стимул. Привыкание в той или иной степени присутствует в разных рефлексах. Степень его выраженности зависит от специфичности стимула, силы раздражения и состояния организма. В самом общем виде привыкание — это постепенное снижение реакции в ходе повторений стимула. Отличие привыкания от утомления и истощения в том, что реакцию можно восстановить простым изменением стимула. Наиболее отчетливо привыкание выражено в системе ориентировочного рефлекса.