Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Физико-химические гомеостатические механизмыСодержание книги
Поиск на нашем сайте
К физико-химическим механизмам кислотно-основного гомеостазиса относятся буферные системы внутренней среды организма и тканевые гомеостатические обменные процессы. 15.2.1. Буферные системы внутренней среды организма Основными буферными системами внутриклеточной, межклеточной жидкости и крови являются бикарбонатная, фосфатная и белковая буферная система, причем из последней для крови особо выделяют гемоглобиноный буфер. Основное значение для поддержания pH межклеточной жидкости и плазмы крови имеет бикарбонатная буферная система. Угольная кислота в плазме и межклеточной жидкости присутствует в четырех формах: физиче КН2СОг = Анион НСО3 является общим как для кислоты, так и для соли, а соль диссоциирует сильнее, поэтому данный анион, образуясь из бикарбоната, будет подавлять диссоциацию угольной кислоты, т. е. практически все количество аниона НСО7 в бикарбонатном буфере происходит из NaHCO3 Следовательно: „„ [}Г][НСО'3] К[Н2СО3] КН2СО3 = [Я2Си3] [NaHCO3] (формула Гендерсона, где К —константа диссоциации угольной кислоты). В связи с использованием отрицательного логарифма концентрации формула, названная уравнением Гендерсона—Гассельбаха, для бикарбонатного буфера приняла выражение:
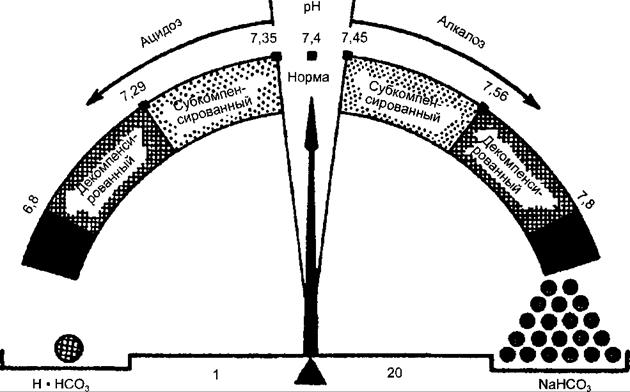
При физиологических значениях pH отношение концентрации углекислоты к бикарбонату составляет 1:20 (рис. 15.1). В условиях взаимодействия бикарбонатного буфера с кислотами происходит их нейтрализация с образованием слабой угольной кислоты. Углекислый газ, появляющийся при ее разложении, удаляется через легкие. Избыток оснований, взаимодействуя с бикарбонатным буфером, связывается с угольной кислотой и приводит в конечном счете к образованию бикарбоната, излишки которого удаляются, в свою очередь, из крови через почки. Другой буферной системой плазмы крови, а в большей мере внутриклеточной среды, является фосфатная буферная система. Она образована одно- и двузамещенными слоями фосфорной кислоты, где однозамещенные соли являются слабыми кислотами, а двузамещенные — имеют щелочные свойства. Уравнение для фосфатного буфера следующее:
Двухосновной фосфатной соли содержится в плазме в 4 раза больше, чем кислой одноосновной. Общим анионом в этой системе является НРО4. Ее буферная емкость меньше, чем бикарбонатной, так как и фосфатов в крови содержится меньше, чем бикарбонатов. Принцип действия фосфатного буфера аналогичен бикарбонатному, хотя роль его в плазме крови мала и, в основном, сводится к поддержанию концентрации бикарбоната при реакции буфера с избытком угольной кислоты. В то же время в клетках значение фосфатного буфера велико. Третьей буферной системой клеток, плазмы крови и межклеточной жидкости является белковая. Белки выполняют буферную роль из-за их амфо-
Рис. 15.1. Кислотно-основное равновесие. Стрелка весов указывает на значения pH в норме и при смещениях в сторону ацидоза или алкалоза. На чашах весов помещены Н2СО3 (дыхательная компонента) и NaCO3 (недыхательная компонента), что отражает соотношение между ними в норме 1:20.
терности, а характер их диссоциации зависит от природы белка и актуальной реакции внутренней среды. При этом глобулины обладают более выраженной кислой диссоциацией, т. е. отщепляют больше протонов, чем гидроксильных ионов, и соответственно играют большую роль в нейтрализации щелочей. Белки, содержащие много диаминокислот, диссоциируют больше как щелочи и поэтому в большей мере нейтрализуют кислоты. Буферная емкость белков плазмы крови сравнительно с бикарбонатной системой невелика, однако в клетках тканей ее роль может быть очень высокой. Наибольшую буферную емкость крови обеспечивает гемоглобиновая буферная система. Содержащаяся в гемоглобине человека аминокислота гистидин (до 8,1 %) имеет в своей структуре как кислые (СООН), так и основные (NH2) группы. Константа диссоциации у гемоглобина ниже, чем pH крови, поэтому гемоглобин диссоциирует как кислота. Оксигемоглобин является более сильной кислотой, чем редуцированный гемоглобин. При диссоциации оксигемоглобина в капиллярах тканей с отдачей кислорода появляется большее количество щелочно-реагирующих солей гемоглобина, способных связывать Н-ионы, поступающие из кислот тканевой жидкости, например угольной кислоты. Оксигемоглобин обычно представляет собой калиевую соль. При взаимодействии кислот с калиевой солью оксигемоглобина образуется соответствующая калиевая соль кислоты и свободный гемоглобин со свойствами очень слабой кислоты. Гемоглобин в тканевых капиллярах связывает углекислый газ за счет аминогрупп, образуя карбогемоглобин: Hb- NH2+CO2 -> Hb- NHCOOH. Таблица 15.3. Относительная эффективность буферных систем крови
Для кислотно-основного гомеостазиса важное значение имеет также обмен анионов СГ и НСО7 между плазмой и эритроцитами. Если в плазме крови увеличивается концентрация углекислоты, то образующийся при диссоциации NaCl анион С1 входит в эритроциты, где образует КС1, а ион Na+, для которого мембрана эритроцита непроницаема, соединяется с избытком НСО7, образуя бикарбонат натрия, восполняя его убыль в бикарбонатом буфере. При снижении концентрации углекислоты в бикарбонатом буфере происходит обратный процесс — анионы СГ выходят из эритроцитов и соединяются с избытком Na+, освободившимся из бикарбоната, этим, следовательно, предотвращается ощелачивание плазмы. Буферные системы плазмы крови и эритроцитов имеют разную относительную эффективность. Так, эффективность буферных систем эритроцитов выше (за счет гемоглобинового буфера), чем плазмы крови (табл. 15.3). Концентрация Н-ионов уменьшается в направлении: клетка — межклеточная среда — кровь. Это свидетельствует о том, что наибольшую буферную емкость имеет кровь, а наименьшую — внутриклеточная среда. Образуемые в клетках при метаболизме кислоты поступают в межклеточную жидкость тем легче, чем больше их образуется в клетках, так как избыток Н-ионов повышает проницаемость клеточной мембраны. В буферных свойствах межклеточной среды играет роль соединительная ткань, особенно коллагеновые волокна, известные как «ацидофильные». На минимальное накопление кислот они реагируют набуханием, быстро поглощая кислоту и освобождая от Н-ионов межклеточную жидкость. Эта способность коллагена объясняется свойством абсорбции. 15.2,2. Тканевые гомеостатические обменные процессы Кислотно-основное состояние поддерживается в пределах физиологических значений pH и путем метаболических превращений в тканях. Это достигается за счет совокупности биохимических и физико-химических процессов, которые обеспечивают: 1) потерю кислотных и щелочных свойств продуктов обмена веществ, 2) их связывание в средах, препятствующих диссоциации, 3) образование новых, более легко нейтрализуемых и выводимых из организма соединений. Так, например, органические кислоты могут соединяться с продуктами белкового обмена (бензойная кислота с глицином) и тем самым терять кислые свойства. Избыток молочной кислоты ресинтезируется в гликоген, кетоновых тел — в высшие жирные кислоты и жиры. Неорганические кислоты нейтрализуются солями калия, натрия, освобождающимися при дезаминировании аминокислот аммиаком, образующим аммонийные соли. После внутривенного введения животным кислоты 43 % ее количества нейтрализуется бикарбонатом плазмы крови, 36 % нейтрализуются за счет клеточного натрия, а 15 % — за счет выходящего из клеток калия. Основания нейтрализуются преимущественно молочной кислотой, образуемой из гликогена при ощелачивании микроокружения клеток. В поддержании внутриклеточного pH существенную играет роль обмен производных имидазола и его изомера пиразола. Особенности пятичленного кольца этих соединений определяют их амфотерные свойства, т. е. способность быть одновременно и донатором, и акцептором протонов. Ими- задол способен очень быстро образовывать соли с сильными кислотами и щелочными металлами. Наиболее распространенным соединением имидазола является а-аминокислота гистидин, участвующая в кислотном и основном катализе. Сильные кислоты и щелочи могут растворяться в липидах, обладающих низкой диэлектрической константой, что препятствует их диссоциации. Наконец, органические кислоты могут подвергаться окислению с образованием летучей слабой угольной кислоты.
|
||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-09; просмотров: 163; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.115 (0.009 с.) |