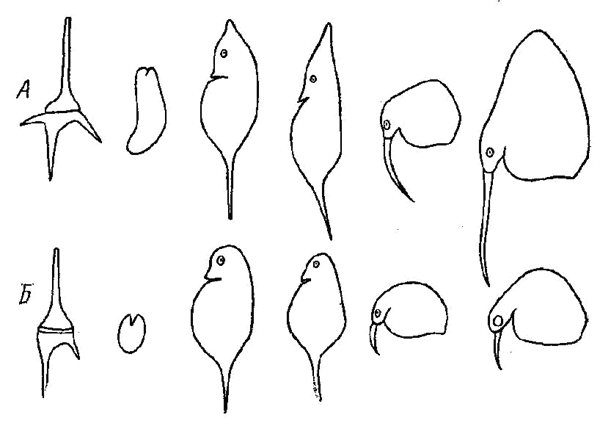Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Краткий очерк истории понятий
Сегодня мы будем рассматривать жизненные формы гидро-бионтов, обитающих в толще воды вне связи с субстратом как опорным элементом, в том числе и пленкой поверхностного натя-жения, в течение всей активной жизни или на определенных ста-диях жизненного цикла.
Как обычно, проследим краткую историю возникновения по-нятия и термина «планктон» и области исследований сообществ организмов, парящих в толще воды.
Еще в 1843 г. на о-ве Гельголанд И. Мюллер изучал «чудесный пелагический мир» Северного моря, пользуясь планктонной се-тью. Позднее датский натуралист П.Э. Мюллер (1870) обнаружил,что прозрачные воды швейцарских озер не безжизненны, а населе-ны характерным и специфическим пелагическим сообществом.
Термин «планктон» введен в науку впервые немецким физио-логом В. Гензеном в 1887 г. (Hensen, 1887) и произведен от грече-ского слова «planktos», что означает «блуждающий вокруг». Но-вый термин вскоре был принят Эрнстом Геккелем (Haeckel, 1890), который от него произвел прилагательное «планктонный» («planktonisch»), а для научной дисциплины, которая занимается этой областью биологии, предложил наименование «планктоло-гия» («Planktologie»).
Планктонология и гидробиология долгое время существовали параллельно: старейший гидробиологический журнал в первые годы (1906 – 1922) носил название «Архив гидробиологии и планктоноведения» («Агchiv fur Hydrobiologie und Plankton-kunde»).Выход в свет первого научного журнала ряд ученых счи-тают также годом основания гидробиологии.
В 1906 г. была опубликована книга С. Апштейна «Планктон пресных вод». Примененная Апштейном удобная для пресных вод модель планктонной сети используется и по сей день под назва-нием «сеть Апштейна».
Состав зоопланктона и экологические группы
В 1903 г. методом центрифугирования воды Г. Ломан от-крыл мельчайших представителей планктона, названных в 1909 г.
нанопланктоном, или(наннопланктоном) - карликовым. В 1910 году С. Везенберг-Лунд детально проследил цикло-морфоз,свойственный многим представителям пресноводногопланктона, особенно некоторым ракообразным (дафнии, босми-ны) и коловраткам (см. Лекция 6), обнаруживающих закономер-ные сезонные изменения формы тела. Буркхардт (Burckhardt, 1920) предложил термины «планктический» (нем. «planktisch» и «планктер» («Plankter», «planktor»)).
Виктор Гензен под планктоном понимал «все, что в воде носи-мо, наверху или в глубине, мертвое или живое. Решающим при
этом является то, носимо ли животное пассивно с водой или оно со-храняет некоторую степень самостоятельности против этой силы дрейфа? Поэтому рыбы принадлежат к планктону самое большее в стадии икры и молоди, но не взрослого животного (нектон); копе-поды хотя и плавают энергично, однако пассивно увлекаются водой
и поэтому должны быть причислены к планктону» (Hensen, 1887; цит. по: Винберг ГГ, 1975, см. лекцию 7). В.М. Рылов (1922) под планктоном предлагает понимать «со-вокупность организмов, обитающих в толще воды вне связи с субстратом как точкой опоры и не способных противостоять даже слабому течению вследствие отсутствия или слишком малой силы их активных движений».
Для обозначения совокупности всех взвешенных в воде эле-ментов, независимо от их происхождения, как живых, так и нежи-вых, Р. Колквитц (R. Kolkwitz 1912) предложил термин «сестон». К нему принадлежат как живые (биосестон), так и неживые эле-менты (абиосестон). Планктон представляет собою, следователь-но, часть биосестона.
Биосестон включает в себя четыре сообщества организмов: нейстон и плейстон, планктон и нектон. Что касается абиосестона, то входящие в его состав нерастворимые в воде мертвые взвеси обычно именуются терминами «детрит» и «триптон». Последний термин был предложен Вильгельми (Wilhelmi, 1916).
В составе планктона можно встретить представителей всех царств и систематических групп: бактерии (бактериопланктон), расте-ния – водоросли (фитопланктон), животные (зоопланктон). Зоо-планктон составляют обычно три систематические группы беспо-звоночных: коловратки (Rotatoria, класс), ветвистоусые раки (Cladocera, отряд), веслоногие раки (Copepoda, отряд). Большин-ство каляноид (Calanoida, Copepoda) в течение всей жизни, кроме стадии покоящихся яиц, ведет планктонный образ жизни; цикло-пы (Cyclopida, Copepoda) населяют и водную толщу, и являются компонентом микрозообентоса; гарпактициды (Harpacticoida, Copepoda) считаются исключительно бентическими животными, но достаточно часто встречаются в планктоне.
Рис. 1. Представители морского меропланктона (по Одуму, 1975): 1 – личинка червя Platynereis, 2 – зоеа краба Emerita, 3 – личинка мшанки, 4 – личинка сессильных оболочников, 5 – пилидий немертин, 6 – плутеус морского ежа, 7 – икринка рыбы, 8 – трохофора полихет, 9 – велигер моллюсков, 10 – плутеус морской звезды, 11 науплиус усоногих, 12 – циприсовая личинка усоногих, 13 – планула кишечнополостных, 14 – медузоидная стадия гидроидов
По размерному признаку различают мегало-, макро-, мезо-, микро-, нанно- и пикопланктон, к которым соответственно относят-ся организмы крупнее 5 см (медузы, до 100 см), около 5 мм (круп-ные рачки), 0,5 мм (коловратки, мелкие рачки), 50 мкм (простей-шие – растительные жгутиконосцы, мелкие коловратки), 5 мкм (водоросли, бесцветные жгутиконосцы) и менее 5 мкм (мелкие во-доросли, бактерии). По степени привязанности организмов к вод-ной толще различают голо- и меропланктон. К первому принадле-жат организмы, которые всю активную жизнь проводят в толще воды, и только покоящиеся стадии (почки, яйца и др.) могут нахо-диться на дне. К меропланктону (рис. 1) относятся формы, обитаю-щие в толще воды только на каком-то отрезке своего активного су-ществования, а остальную часть жизни ведущие иной образ жизни (пелагические личинки донных животных, икра и личинки рыб, ли-чинки плейстонта – физалии). Есть организмы, которые чередуют свободное плавание в толще воды с моментами прикрепления к твердому субстрату, как это имеет место у некоторых коловраток, снабженных ногой (Brachionus и др.), у эвгленовых (Colacium cyclopicola),инфузорий(Trichodina pediculus, Stentor)и некоторыхрачков сем. Сhydoridae и др. Эти организмы могут быть названы бенто-планктическими, или факультативно-планктическими.
В состоянии свободного плавания или парения протекает вся активная жизнь голопланктона. Только их стадии покоя (напри-мер, покоящиеся яйца планктонных коловраток и ракообразных) опускаются на дно водоема, где и пребывают некоторое время в покоящемся, неактивном состоянии (рис. 2 – 4).
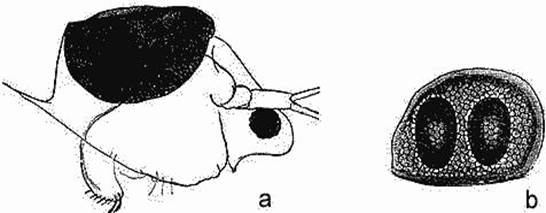
Рис. 2. a. Седлышко гипонейстонного рачка Scapholeberis mucronata в мертвой раковине. b. Седлышко Moina rectirostris
Рис. 3. Зимние яица. a. Sida cristallina, b. Diafanosoma brachyura, c. Polyphemus pediculus
Рис. 4. a. Раковинка Eurycercus lamellatus. b. Зимнее яицо Alona qyadrangularis
Образованию таких стадий в огромном большинстве случаев предшествует половой процесс (оплодотворение). Однако раз-множение очень многих планктонтов происходит или путем про-стого деления, или партеногенетически, – партеногенез свойствен планктону в широкой степени. В том и другом случае размноже-ние и развитие имеет место в самом планктоне.
Размножение простейших животных (Protozoa) и водорослей планктона посредством деления происходит в состоянии плавания или парения, причем те и другие продолжают свое планктонное существование.
Некоторые организмы (ветвистоусые ракообразные Clado-cera)вынашивают партеногенетические яйца в своем теле–вособой выводковой камере, где и протекает их развитие, причем рачок продолжает вести планктонный образ жизни.
Рис. 5. Simocephalus vetulus с Рис. 6. Limnosida frontosa c яйцами в выводковой камере зародышами в выводковой камере
Это же следует сказать и о планктонных коловратках (Rotatoria), прикрепляющих такие яйца к заднему концу тела и тас-кающих их за собой, пока из них не вылупится молодое животное, есть также коловратки, откладывающие партеногенетические яйца прямо в воду, и развитие последних протекает во взвешенном в во- де состоянии, без опускания яйца на дно водоема, а также коловратки, прикрепляющие яйца к планктон-ным водорослям.
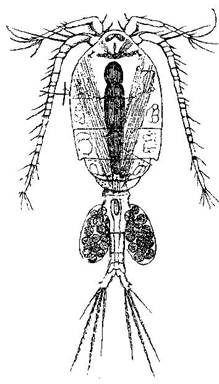
В состав планктона входят также организмы, размножающие-ся исключительно половым путем. В пресноводном планктоне очень крупную роль играют веcлоногие ракообразные (Copepoda (рис. 7)), у которых партеногенез вообще от-сутствует. Эти рачки образуют яй-ца двоякого рода – покоящиеся и непокоящиеся. Последние разви-ваются быстро, тогда как первые некоторое, довольно продолжи-тельное время остаются в стадии покоя, как бы застывая на ранних стадиях своего развития. При этом только покоящиеся яйца опускают- ся на дно, где и лежат, пока из них не выйдет личинка (науплиус), сразу же приступающая к планктонной жизни.
Напротив, все развитие непокоящихся яиц протекает в связи с рачком, к которому эти яйца прикрепляются в одном или двух яй-цевых мешках, причем вынашивающий яйца рачок продолжает вести планктонный образ жизни. Из этих яиц вскоре вылупляется личинка, науплиус, дальнейшее развитие которой протекает в со-стоянии свободного плавания.
В таком же состоянии происходит и питание планктонных ор-ганизмов. Они находят себе пищу в достаточном количестве в толще воды или в виде других планктонтов, или в виде взвешен-ных в воде мельчайших частичек органического происхождения, или, наконец, в виде растворенных в воде минеральных и органи-ческих веществ.
Таким образом, важнейшие жизненные функции планктонных организмов, как питание, развитие и размножение, протекают в плавающем или парящем в толще воды состоянии, независимо от какого-либо иного опорного элемента, кроме самой водной среды. Планктонный образ жизни определяет организацию планктонтов
и все их жизненные проявления, тесно связанные со специфиче-скими условиями местообитания.
Теория парения
Первым, кто пытался объяснить сезонные и локальные изме-нения формы у планктических организмов, был Везенберг-Лунд (Wesenberg- Lund, 1900). Парение планктонтов тесно связано с плотностью и вязкостью воды. В холодной плотной воде парение обеспечивается минимальными отношениями объема и поверхно-сти тел. С уменьшением плотности при повышении температуры воды парение должно обеспечиваться увеличением удельной по-верхности тела. При этом организмы становятся длиннее, тоньше, уплощаются и приобретают или увеличивают выросты тела. При этом изменяется архитектоника тела. Организмы так сильно могут менять форму тела, что долгое время специалисты описывали но-вые для науки виды в зимнее и летнее время. Позже, после на-блюдений Везенберг-Лунда, это явление назвали цикломорфозом. Но не только изменением формы обеспечивается парение, но и
физиологическими характеристиками: увеличение содержания жира, газовых камер в цитоплазме, уменьшающих удельный вес организмов относительно удельного веса воды.
Оствальд (Ostwald, 1902) в результате своих эксперименталь-ных исследований показал, что гораздо большее значение, чем из-менение удельного веса воды, должно иметь изменение в связи с изменением температуры и вязкости, или внутреннего трения воды. Оствальд предложил формулу, в которой установлена зави-симость скорости погружения (флотация) (а) от удельного веса организмов относительно воды (b – остаточная масса – разница между массами организма и вытесненной им воды), вязкости воды (с) и сопротивление формы (d): a = b
c × d
Снижение остаточной массы (b)
Плотность воды заметно возрастает с понижением температу-ры, а также с повышением солености и давления, в связи с чем ус-ловия плавучести организмов заметно меняются. В соответствии с этим плотность планктонтов регулируется так, что она приближа-ется к плотности воды. Как правило, плотность пресноводных планктонных организмов не превышает 1,01 – 1,02 г/см3, мор-ских – 1,03 – 1,06 г/см3, и их плавучесть близка к нейтральной. Если гидробионты совершают вертикальные миграции или пере-мещаются в участки с иной плотностью воды, они обычно изме-няют свою плотность, модулируя состав тела. Плотность его от-дельных компонентов выражается следующими средними величинами (при 20°С): сквален – 0,86 г/см3; восковые эфиры – 0,90, диацилглицериновый эфир – 0,91; триацил-глицерин – 0,92, белок – 1,33, скелетная ткань – 2 – 3 г/см3.
Снижение остаточной массы может достигаться уменьшением количества костной ткани, белка в тканях, заменой тяжелых солей более легкими, отложением большого количества жира, заменой более плотного жира менее плотным, образованием полостей, на-полненных воздухом. Редукция тяжелых скелетных образований хорошо прослеживается у пелагических моллюсков. Так голожа-берные и крылоногие моллюски в морском планктоне вовсе не
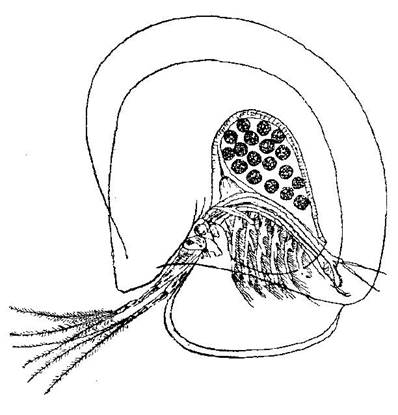
имеют скелета-раковины. Крупная кладосера Leptodora kindtii утра-тила раковину и парит в воде, скачкообразно передвигаясь за счет сильных антенн. Рачки сем. Polyphemidae практически не имеют раковины. Карапакс Copepoda представлен тонкой кутикулой. Некоторые планктонные организмы выделяют стекловидно-прозрачную студенистую массу, облекающую тело снаружи. Это наблюдается, например, у синезеленых водорослей Аnabaena и Мicrocystis,у целого ряда зеленых водорослей,а также и у планк-тонных животных, например, у коловраток. Благодаря очень незна-чительному удельному весу таких богатых водою оболочек, обле-каясь ими, организмы выигрывают в смысле повышения способности поддерживаться во взвешенном состоянии. Рассматри-ваемые образования имеют одновременно и другие назначения, в частности служат защитным приспособлением. Так, например, бла-годаря объемистой студенистой капсуле, облекающей тело Holopedium,этот ветвистоусый рачок не заглатывается рыбами(рис. 8).
Рис. 8. Holopedium gibberum Zadd. (По Лиллиеборгу)
Один из самых распространенных способов снижения оста-точной массы – повышение содержания воды в теле. Ее количест-
во у некоторых сальп, гидромедуз, гребневиков Cestus veneris, сцифоидных медуз (Aurelia aurita) и трахимедуз Сагmarina дости-гает 99%. При таком содержании воды остаточная масса организ-ма приближается к 0, и способность к пассивному флотированию становится практически безграничной. Наиболее обычный способ снижения плотности у гидробио-нтов – накопление жира. Богаты им ночесветки Noctiluca, радио-лярии Spumellaria, ветвистоусые и веслоногие рачки. Жировые капли имеются в пелагической икре ряда рыб (кефалевые, камба-лы, скумбрия). Жир вместо тяжелого крахмала в качестве запас-ного питательного вещества отлагается у планктонных, диатомо-вых и зеленых водорослей.
Эффективное средство повышения плавучести – газовые включения в цитоплазме или специальные воздушные полости. Газовые вакуоли есть у многих планктонных водорослей. В про-цессе эволюции накопление газовых пузырей на талломах Sargas-sum natans иS. fleutans превратило эти бурые водоросли из дон-ных в гипонейстонные формы. Так, в протоплазме планктонных корненожек образуются содержащие углекислоту вакуоли. Благо-даря их присутствию вес животного понижается. Особенно заме-чательна в этом отношении личинка Corethra, ведущая планктон-ный образ жизни. Внутри полости ее тела находится две пары воздушных пузырей, представляющих видоизменение трахейной системы, которая у этой личинки вполне замкнута. Пузыри на-полнены газом, выделяемым самой личинкой. Передняя пара пу-зырей лежит около головы, задняя – у хвостового плавника. Пу-зыри служат личинке в качестве прекрасного гидростатического аппарата; благодаря их попарному расположению спереди и сза-ди, животное держится в воде горизонтально.
Газовый пузырек имеют в своей цитоплазме раковинные аме-бы Difflugia hydrostatica и Arcella. Аналогичные плавательные приспособления особенно характерны у некоторых морских планктонных животных (плавательный пузырь у сифонофор).
Сопротивление формы (d)
В этом направлении у планктонных организмов также наблю-дается ряд приспособлений. Тенденция к возможно большему увеличению поверхности тела при его возможно наименьшем
объеме – явление в общем очень характерное для планктонных организмов.
Целому ряду планктонтов свойственно удлинение морфоло-гической продольной оси организма. Так, например, туловище и брюшко Leptodora kindtii имеет вид весьма улиненного цилиндра; равным образом удлинен и передний, головной отдел этого рачка.
Нужно заметить, что образование игловидных придатков – явление очень частое среди планктонных животных. Так, ветви-стоусый рачок Вythоtrephes сзади несет весьма длинную тонкую иглу (придаток постабдомена). При плавании животного эта игла направлена горизонтально – обстоятельство весьма существенное, так как только при таком положении она может иметь назначение
в качестве приспособления для повышения сопротивления формы. У Bythotrephes рассматриваемая игла одновременно служит рулем и органом, сохраняющим устойчивость тела при плавании. Ана-логичные придатки мы находим у планктонной коловратки
Notholсa longispina.
Вязкость воды (с)
При 25°С вязкость воды уменьшается вдвое по сравнению с тем, что имеет место в воде при 0°С, это означает, что организм, для того чтобы замедлить свое погружение и держаться во взве-шенном состоянии в том же слое воды в летней воде, он должен увеличить удельную поверхность своего тела и увеличить тем са-мым сопротивление формы или уменьшить размеры тела.
Так как все изменения формы направлены к тому, чтобы уве-личить летним животным их относительную поверхность (d) пу-тем уменьшения их размеров, путем удлинения их продольной оси или образования разнообразных выростов, понижающих ско-рость их погружения, то, естественно, сезонные вариации должны рассматриваться как приспособления к меняющейся вязкости во-ды – цикломорфоз (рис. 9).
При отсутствии температурных вариаций в водоемах теплых и северных регионов и альпийских озерах не наблюдается цикло-морфных явлений. Наиболее отчетливо они наблюдаются у круг-логодичных видов, переживающих относительно низкие зимние и высокие летние температуры, например, Daphnia cucullata, Bosmi-na coregoni,коловратки Keratella quadrata и др.
Рис. 9. Цикломорфоз планктонных организмов (по Зернову, 1949): А –
летник формы (слева направо): Cratium hirundinella, Asplancha priodonta, Daphnia hyaline, Hyalodaphnia cuculata, две расы Bosmina coregoni; Б –зимние формы тех же видов
В широтном плане явления цикломорфоза просматриваются на экологических эквивалентах из северных и южных морей.
Изложенная выше теория парения Везенберг-Лунда и Ост-вальда и толкование с ее позиций сезонных вариаций развились в одно из центральных знаний классической планктологии. В даль-нейшем явление сезонных вариаций продолжало привлекать вни-мание исследователей и правильность этой теории была подтвер-
ждена на разных объектах (Ceratium, Phyliopoda, Keratella). Однако отмеченные явления цикломорфоза не носят абсо-лютного характера. Есть масса примеров, противоречащих теории парения Везенберг-Лунда и Оствальда. Их связывают с периодом размножения, питанием, способом размножения, полом и номе-ром партеногенетических поколений, гидродинамикой (в прибре-жье крупнее, чем в открытых участках водоема, подверженных частому ветровому перемешиванию) и т.д.
Вертикальные миграции зоопланктона
Организмы зоопланктона не могут противостоять сносу, но в толще воды, даже по ходу сноса слабым течением они постоянно перемещаются (мигрируют) по вертикали, находя свое место в температурном градиенте, уходят вглубь от яркого света, находят пищу растительную, микробную или, хищничая, одновременно спасаясь от врагов, выбирают условия для размножения, зимовки
и т.д. Исследуя вертикальное распределение зоопланктона или от-дельных систематических групп в течение суток в разные сезоны года, ученые видят определенные закономерности в этом распре-делении, отражающие биологические особенности отдельных ви-дов (питание, размножение, онтогенез), популяционные отноше-ния видов, общую экологическую ситуацию в водоеме и изменение ее под влиянием хозяйственной деятельности человека: загрязнения, подогрев или изменения гидрологического режима. Мы рассмотрим ряд миграций зоопланктеров, отражающих се-зонные и суточные изменения сред обитания и биологические особенности видов.
Сезонные миграции
Сезонные изменения вертикального распределения планктона изучаются по биологическим сезонам: зима, весна, лето и осень. Наиболее отчетливо они выражены в высоких и умеренных широ-тах, а также в глубоких водоемах.
Различают зимние, летние и круглогодичные виды зоопланк-тона. Весенний и осенний зоопланктон обычно составляют онто-генетические стадии первых, вторых и третьих.
Сезонные явления в распределении зоопланктона можно про-следить на примере оз. Плещеево (Столбунова, 2006).
Горизонтальное распределение зоопланктеров в озере далеко неравномерно и определяется комплексом факторов: глубиной, динамикой водных масс, температурой. Важную роль играет пи-щевой фактор и даже грунты, на поверхности которых зимуют не-которые виды циклопид, покоящиеся яйца кладоцер, коловраток.
В зимний период, когда влияние гидродинамических факто-ров на водную массу ослабляется, зоопланктон распределен осо-бенно неравномерно. Его максимальное развитие наблюдается в
марте. Наиболее богата центральная часть озера и участки с глу-бинами до 7 м, где летом располагается эпилимнион (до 1,05– 1,57 г/м3). Основу зоопланктона составляют холодолюбивые ко-ловратки и Eudiaptomus graciloides.
После вскрытия водоема прогрев мелководий от 7,5°С до 13,4°С вызывал массовое развитие коловраток до 70 тыс. экз./м3.
В глубоководных, менее прогретых участках с температурой воды у поверхности 6,8 – 8,1°С развивался в основном холодолюбивый Cyclops kolensis (до135тыс.экз./м3и2,51г/м3). С наступлением лета, когда температура воды у поверхности достигает 21 – 24°С, возрастает роль кладоцер и летних веслоно-гих. В поверхностном слое (0 – 2 м) в центре озера Daphnia cucul-lata достигала2,22г/м3, Bosmina coregoni –до1,95г/м3, Mesocyc-lops leuckarti –до0,50г/м3.В глубоководной зоне озера,а такжена свале глубин образуются наиболее плотные скопления зоо-планктона (биомасса до 5 г/м3 и более). При любом из преобла-дающих направлений ветра наблюдается тенденция к увеличению биомассы зоопланктеров в двух вихревых зонах циркуляции вод в озере и разрежения плотности между ними (рис. 10).
Рис. 10. Схема интегральной циркуляции вод (а, по: Поддубный, Литвинов, 1983) и горизонтальное распределение зоопланктона (б)
в слое 0–2 м в летнее время (из: В.Н. Столбунова, 2006) В осенний период с понижением температуры воды до 11 – 13°С в планктоне еще присутствуют летние Keratella quadrata, K. cochlearis, Conochilus unicornis,из ракообразных многочисленна
Bosmina coregoni,встречаются Daphnia cucullata и Eudiaptomus graciloides. Распределение зоопланктона в поверхностном слоеопределяется погодными условиями, и при ветровом перемеши-вании воды оно более равномерно.
Межгодовые колебания общей численности зоопланктона в поверхностном слое пелагиали озера за вегетационный период мало отражались на величине биомассы, так как основу последней составляли ракообразные, за исключением 1989 г., когда преобла-дала крупная Asplanchna priodonta (табл. 1).
Таблица 1
Средние количественные показатели зоопланктона
в поверхностном слое 0 – 2 м профундали оз. Плещеево за вегетационный период в разные годы
Вертикальное распределение зоопланктона в озере изменя-ется в зависимости от условий сезона года, прежде всего от тер-мического режима и связанной с ним стратификации водной мас-сы, а также от немаловажного пищевого фактора.
В зимний период при устойчивой стратификации водной тол-щи и температуре воды 0 – 2,9°С, в поверхностном слое наиболее многочисленны коловратки родов Keratella, Polyarthra, Synchaeta;
здесь максимальное содержание кислорода, минимальная тем-пература, присутствуют бактерии и водоросли. Придонные ско- пления образуют Filinia maior и Conochiloides natans, хорошо пе- реносящие дефицит кислорода, а также Asplanchna priodonta. В некоторые годы придонные скопления образовывал эвритермный Conochilus unicornis. Eudiaptomus graciloides заселяет всю толщупелагиали озера с тенденцией возрастания численности к поверх-ности. Его количество значительно больше, чем Cyclops kolensis, который образует небольшие скопления в придонных слоях. Вет-
вистоусые Bosmina coregoni, Daphnia longispina и D. cristata сосре- доточены в металимнионе и придонных слоях.
Весной, в период гомотермии, при температуре 5–6°С коло-вратки и ракообразные распределяются по вертикали относитель-но равномерно. В конце мая, когда температура воды повышается до 17,0 – 17,5°С, начинает формироваться слой температурного скачка. В период летней стратификации водной толщи вертикаль-ное распределение популяций зоопланктона позволяет выделить два основных комплекса: тепловодный многокомпонентный (Ke-ratella quadrata, K. cochlearis, Pompholyx sulcata, Conochilus un-icornis, Daphnia cucullata, Bosmina coregoni, Eudiaptomus graci-loides и др.)и холодноводный малокомпонентный(Keratella hiemalis, Filinia maior, Daphnia cristata).
Наибольшей плотности зоопланктон достигает в эпилимнио-не, где располагается оксиклин и интенсивно идут первичные продукционные процессы. По мере охлаждения озера, ускорения циркуляции вод и значительном ветровом перемешивании водной толщи наступает осенняя гомотермия. Распределение коловраток и ракообразных по вертикали становится более равномерным.
Суточные миграции
Суточные миграции зоопланктеров – интереснейшая научная
и практическая тема исследований. Ни одна серьёзная работа по зоопланктону, например диссертация, не обходит эту тему. Здесь изучаются разные стороны биологии и экологии видов, трофиче-ские связи и отношения популяций. Суточные миграции происхо-дят во всем Мировом океане и практически во всех озерах и водо-хранилищах. Трудно себе представить, но это происходит, что триллионы тонн планктонных организмов каждые сутки подни-маются из глубин к поверхности, питаются там, размножаются и опускаются обратно. Суточные миграции могут быть горизон-тальными и вертикальными.
Примерами горизонтальных миграций могут быть суточные перемещения Polyphemus pediculus из разрозненных групп в ноч-ное время в локальные плотные скопления по мере увеличения освещенности с восходом солнца. К вечеру скопления рассредо-тачиваются. Подобные скопления можно наблюдать у Daphnia magna,но в затененных водоемах в дневное время.При яркомдневном освещении дафнии уходят на глубину.
Изучение вертикальных миграций зоопланктона предполагает экологическую оценку этого грандиозного явления, вероятно дей-ствующего со времени зарождения жизни в толще вод океана. Зада-чи изучения в основном сводятся к установлению причин, времени
и глубины нахождения видов, групп (Cladocera, Copepoda, Rotatoria)
и их представителей на определенных стадиях развития и размно-жения. Наблюдения (отбор проб) проводятся круглосуточно, через 2 – 3 часа, на глубинах от поверхности до дна, обычно через 2 м. Наиболее полные исследования в этом плане проводятся во все 4 биологических сезона и обычно в открытой, глубоководной части озера или водохранилища. В морях это обычно специальные эпизо-дические исследования из-за их трудоемкости и дороговизны. Существует целая история изучения вертикальных миграций. Первые исследования суточных вертикальных миграций ра-кообразных были проведены на оз. Леман в 1874 г. (Forel, 1874, 1879) и на оз. Боденском (Weismann, 1877), где наблюдались су-
точные миграции Leptodora hyalina (L. kindtii). Уже к началу ХХ в. были установлены некоторые закономер-ности суточных вертикальных миграций зоопланктона. Эти зако-номерности касались интенсивности и размаха (амплитуды) ми-граций у разных видов, неодинакового поведения одних и тех же видов в разных водоемах, изменения характера миграций в связи с возрастом (стадией), с полом, в зависимости от сезона года, изме-нения времени подъема к поверхности и ухода вглубь у разных видов. Делались тогда же и первые попытки истолковать это яв-ление, установить факторы, управляющие им.
Кушинг (Gushing, 1951) изучал миграции ракообразных в све-товом градиенте, когда рачки находили свой световой оптимум, где они могут успешно питаться.
Величина светового оптимума неодинакова у разных видов. В связи с этим время появления зоопланктеров у поверхности и опускания их на глубину у разных видов может быть весьма не-одинаковым. Различают мигрантов сумеречных и ночных. Первые концентрируются у поверхности в большом количестве вечером и на рассвете, вторые – ночью, количество первых у поверхности падает днем и ночью, а у вторых – днем.
Оказалось, мелководные виды ведут себя как послеполуден-ные мигранты, более глубинные – как вечерние, а еще более глу-
боководные – как ночные мигранты. При этом чем глубже вид живет в озере, тем сильнее варьирует его средняя дневная глубина от случая к случаю и тем больше вертикальная протяженность его популяции. Световой оптимум различен у разных стадий, полов и даже генераций одного вида, может зависеть от физиологического состояния организма.
Отсутствие вертикальных суточных миграций отмечено в олиготрофных гумифицированных озерах с коричневой водой и незначительной прозрачностью из-за довольно однородных усло-вий во всей толще воды в течение суток.
Хатчинсон (Hutchinson, 1967) различает три типа ночных ми-граций:
1. Подъем начинается перед или вскоре после захода солнца, и животные достигают поверхности незадолго до полуночи, спуск начинается рано утром, как только небо начинает светлеть (юве-нильные Daphnia longispina в оз. Лох-Дерг, копеподы в оз. Лю-церн, Diaphanosoma в оз. Титизее).
2. Подъем продолжается всю ночь, и максимум у поверхно-сти отмечается перед рассветом или в умеренных широтах ле- том – около 4 ч утра Eudiaptomus gracilis и Eurytemora velox в оз. Люцерн). 3. Нет хорошо выраженного ночного подъема, скорее, имеет место ночное опускание, длящееся всю ночь (Cyclops strenuus в оз. Нозири и Macrohectopus branickii в оз. Байкал). При обзоре поведения родов, видов или одного и того же вида в различные периоды его жизни, в разные сезоны у разных стадий и даже генераций или в разных водоемах обнаруживается удиви-тельное разнообразие суточного цикла с уклонением от общей схемы и даже аномалии в поведении.
Наблюдения показали, что наиболее активно мигрируют взрослые рачки. Молодь (науплии) мигрируют мало и в основном находятся в слое воды, содержащем больше растительного детри-та и разлагающих его микроорганизмов, которые составляют ос-новную пищу науплий и копеподитов. Этим слоем часто оказыва-ется слой температурного скачка.
Онтогенетические миграции
Большая часть пелагических животных, особенно населяю-щих промежуточные и глубинные воды океана, меняют глубину своего обитания на разных стадиях развития. Принципиальная схема возрастных миграций у всех планктонных животных, за не-многими исключениями, однотипна. Половозрелые формы, или во всяком случае половозрелые самки, преобладают обычно на наи-больших глубинах обитания вида и там размножаются. Личинки (или яйца) поднимаются на наименьшие доступные виду глубины. Реже для икрометания поднимаются самки. В обоих случаях ли-чинки откармливаются в богатых пищей верхних слоях и лишь по мере роста опускаются на большие, бедные пищей, но зато менее населенные глубины. Амплитуда возрастных миграций определя-ет степень эврибатности вида и может колебаться от нескольких десятков и сотен до нескольких тысяч метров.
Естественно, указанным образом распределяется не вся попу-ляция, так как у многих глубоководных животных эврибатная мо-лодь встречается по всей глубине обитания вида и лишь преобла-дает в верхних горизонтах; также и половозрелые особи многих видов поднимаются значительно выше глубины своего основного обитания (Бирштейн, 1951; Виноградов, 1955, 1959).
У видов, поднимающихся в поверхностную зону, каждая фаза онтогенетических миграций (во всяком случае в средних и высо-ких широтах) приурочена к определенному времени года, и они в равной степени могут быть названы сезонными миграциями.
Различные генерации одного и того же вида могут мигриро-вать по-разному и в разное время. Среди интерзональных видов, диапазон обитания которых дос-таточно широк, в зависимости от того, где происходит икрометание (в поверхностной зоне или в глубже лежащих водах), можно выде-лить два типа онтогенетических миграций. В одном случае в по-верхностные слои для икрометания поднимаются взрослые формы, в другом – молодь, вылупившаяся в глубинных слоях. В морях Северной Атлантики первым типом миграции обла-дают наиболее многочисленные верхне-, интерзональные виды копепод Calanus finmarchicus (Мантейфель, 1941).
Причины миграций
Более чем столетние исследования миграций зоопланктеров показали несколько разных причин, неодинаковых для водоемов разного типа, широтного расположения, разного светового режи-ма и особенно температурного. Объединяющей причиной может быть нахождение организмами луч
|
|||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-19; просмотров: 532; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.143.244.83 (0.189 с.) |