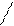Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Нарушение обмена серосодержащих аминокислотСодержание книги
Поиск на нашем сайте
Цистинурия – серосодержащие аминокислоты теряются с мочой в результате нарушение реабсорбции в почках Цистиноз – накопление серосодержащих аминокислот в тканях в результате снижения активности лизосомальных ферментов их распада Гомоцистинурия – с мочой выделяется гомоцистеин в результате нарушения промежуточных стадий этих аминокислот. Гомоцистеин играет роль в патегенезе СС-заболеваний, тромбоза
БИЛЕТ_29 Жировая дистрофия отдельных органов: миокард, печень, болезнь Боткина усиливает отложение ТАГ и показано применение липотропных веществ описаны митохондриальные болезни при которых может страдать бетаокисление жирных кислот. №2 Биосинтез белка (трансляция генов). Сборка полипептидной цепи из составляющих ее АК представляет собой удивительный и очень сложный процесс, который можно представить происходящим в 4 стадии, а именно:1) активация и отбор АК (АТФ-зависимая стадия);2) инициация синтеза полипептидной цепи (ГТФ-зависимая стадия); 3) элонгация полипептидной цепи (ГТФ-зависимая стадия); 4) терминация синтеза полипептидной цепи. (1)– активация и отбор АК. Во всех типах клеток первой стадией трансляции является АТФ-зависимое превращение каждой АК в комплекс: аминоацил-тРНК. Этим достигается две цели: 1) повышается реакционная способность АК в плане образования пептидной связи. 2) АК соединяется со специфической тРНК (то есть происходит отбор). Реакция идет в 2 стадии + Mg++
аминоацил-тРНК-синтетаза - АМФ
аминоацил-тРНК-синтетаза
Аминоацил-тРНК-синтетаза катализирует присоединение аминоацила (аминокислотного остатка) к 3` гидроксильной группе концевого аденозина. Вспомним строение тРНК:
антикодон (2)Инициация – вторая стадия синтеза белков. . Инициаторным кодоном является АУГ, а инициатором выступает метионин-тРНк Считывание происходит в направлении 5` - 3`. Это узнавание требует упорядоченного, идущего с затратой энергии (ГТФ) взаимодействия с диссоциированными рибосомами. Этот процесс происходит с участием дополнительных белков, которые называют факторы инициации (ФИ), их 8. В процессе участвуют 40S и 60S субъединиц рибосом. Рассмотрим подробный механизм инициации. 1) 40S – субъединица рРНК связывается с областью мРНК, предшествующей первому кодону. В этом принимает участие ФИ-3. 2) Первая аминоацил-тРНК, участвующая в трансляции первого кодона, взаимодействует с ГМФ и ФИ-2. Этот образовавшийся комплекс в присутствии ФИ-1 присоединяет тРНК к первому кодону матрицы и образует инициаторный комплекс с 40S субъединицей рибосомы. 3) После высвобождения всех факторов инициации (ФИ-1,2,3) к ГТФ присоединяется 60S субъединица рибосомы, при этом происходит гидролиз ГТФ. Так завершается образование полной 80S-частицы рибосомы. таким образом образуется полный инициаторный комплекс: рибосома – мРНК – тРНК. Схематично весь этот процесс мы можем представить так:1)40S-субъединица рибосомы при участии ФИ-3 присоединяется к нетранслирующей последовательности мРНК непосредственно перед первым кодоном. 2)аминоацил-тРНК, соединяется с ГТФ и ФИ-2 и при участии ФИ-1 присоединяеся к первому кодону, при этом образует с 40S-субъединицей инициаторный комплекс.3)происходит освобождение ФИ-1,2,3.4) 60S-субъединица взаимодействует с ГТФ и затем присоединяется к инициаторному комплексу. Образуется полная 80S-рибосома, имеющая Р-участок и А-участок.5)после образования инициаторного комплекса с первым кодоном, аминоацил-тРНК попадает в формирующийся Р-участок, оставляя А-участок свободным.(3)Элонгация – продолжение синтеза. На этом этапе происходит удлинение пептидной цепи. В полностью сформированной на стадии инициации 80S-рибосома, А-участок свободен. По сути, в процессе элонгации постоянно повторяется цикл из 3 стадий:1) Правильное расположение следующей аминоацил-тРНК.2) образование пептидной связи.3) перемещение новообразованной пептидил-тРНК из А-участка в Р-участок.(1)– присоединение соответствующей (следующей) аминоацил-тРНК в А-участке требует точного узнавания кодона. Это происходит с помощью антикодона тРНК. Присоединение аминоацил-тРНК к рибосоме происходит благодаря образованию комплекса, состоящего из аминоацил-тРНК, ГТФ и белковых факторов элонгации (ФЭ), их тоже несколько. При этом высвобождается комплекс ФЭ – ГДФ и фосфат. Этот комплекс (ФЭ – ГДФ) затем (при участии ГТФ и других белковых факторов) вновь превращается в ФЭ – ГТФ.(2) - альфа аминогруппа новой аминоацил-тРНК в участке А осуществляет нуклеофильную атаку эстерефицированной карбоксильной группы пептидил – тРНК, занимающей Р-участк. Эта реакция катализируется пептидилтрансферазой – белковым компонентом, входящим в состав 60S-субъединицы рибосомы. поскольку АК а аминоацил-тРНК уже активирована, для этой реакции (реакции образования пептидной связи) дополнительной энергии не требуется. В результате реакции растущая полипептидная цепь оказывается прикрепленной к тРНК, находящейся в А-участке. (3) – после удаления пептдильного остатка с тРНК в Р-участки, свободная молекула РНК покидает Р-участок. Комплекс ФЭ-2 – ГТФ участвует в перемещении новообразованной пептидил-тРНК из А-участка в Р-участок, освобождая А-участок для нового цикла элонгации. Совокупность отделения деацилированной тРНК, передвижение новообразованной пептидил-тРНК из А-участка в Р-участок, а так же передвижение мРНК относительно рибосомы, называется транслокацией. трептомицин, связываясь с рибосомными белками, нарушает узнавание кодона антикодоном. Хлоромицитин связывается с А-участком, блокируя элонгацию. Схематично это можно представить так: 1) следующая аминоацил-тРНК благодаря узнаванию с помощью антикодона закрепляется в А-участке. 3) При образовании этой пептидной связи от пептида отделяется тРНК и покидает Р-участок. 4) Новообразованный пептидил-тРНК с помощью комплекса ГТФ – ФЭ2 перемещается из А в Р-участок, а комплекс ГТФ – ФЭ2 гидролизуется до ГДФ – ФЭ-2 и Фк. 5) В результате этого перемещения А-участок освобождается для присоединения новой аминоацил-тРНК. (4)-Терминация – заключительный этап синтеза белка. После многих циклов элонгации, в результате которых синтезируется полипептидная цепь белка, в №3 Классификация жиров. По структуре спирта, лежащего в основе данного липида. 1. Глицерин содержащие жиры: трансацилглицерин, глицерофосфолипиды 2. Сфингосодержашие жиры: сфингофосфолипиды, сфингогликолипиды (цереброзиды, ганглиозиды) 3. Холестеринсодержащие жиры: свободный холестерин, эфиры холестерина, стероиды
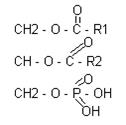
ГЛИЦЕРИНСОДЕРЖАЩИЕ БЕЛКИ: ТАГ – нейтральные жиры. Глицерин содержащие жиры: ФЛ
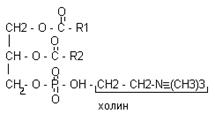
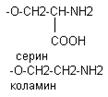
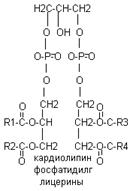
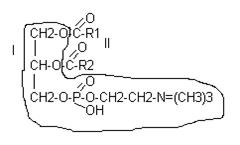
В различных ФЛ фосфатидил содержится с дополнительными N-содержащими веществами: холин, коламин, серин, инозид. Есть особая группа – ацетальфосфатиды (плазмагены). В них в альфа положении нежирная кислота, а альдегид. Фосфатидилглицерины (кардиолипиды).
Фосфатидилхолин-лицетины СФИНГОСОДЕРЖАЩИЕ БЕЛКИ
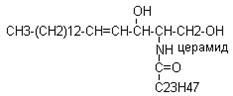
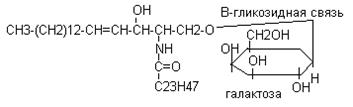
Этот вид липида входят в состав клеточной мембраны в большом количестве в тканях головного мозга, входят в состав сурфактанта легких. В эмбриогенезе их содержание постепенно возрастает в тканях и поэтому содержание в амниотической жидкости характеризует зрелость плода. Гликосфинголипиды: 1. цереброзиды которые содержатся в белом веществе головного мозга, они состоят из церамида и галактозы.
2. Ганглиозиды состоят из церамида и олигосахаридов фрагменты среди которых обычно на концах находятся сиаловые кислоты. Ганглиозиды находятся в сером веществе головного мозга, входят в состав клеточных рецепторов участвуют в связывании некоторых токсинов инородных организмов, механизмы памяти, участие в иммунных реакциях.
ХОЛИСТЕРИНСОДЕРЖАЩИЕ ЖИРЫ 1. Холестерин (содержание 2%) входят в состав клеточной мембраны, является источником синтеза других стероидов организма (желчные кислоты, витамин Д, стероидный гормон). В тканях он находится как в свободном состоянии так и в виде эфиров.
СТРУКТУРА И ФУНКЦИИ КЛЕТОЧНЫХ МЕМБРАН В состав клеточных мембран входят в различных соотношениях белки, жиры и углеводы. На долю белков 50%, липиды 1/3, углеводы 10%.Липиды на 50% представлены фосфолипидами, 1/3 холестерин, остальное сфинголипиды. Углеводы клеточных мембран представлены компонентами гликосфинголипиды, гликопротеиды.Структура клеточной мембраны.В настоящее время принята мозаичная структура клеточной мембраны. Согласно этой модели основу клеточной мембраны составляют фосфолипиды, которые ориентированы в мембране т.о. что гидрофильные участки находятся на поверхности, а гидрофобные в глубине клеточной мембраны. В силу дифильности фосфолипиды образуют билипидный слой. Фосфолипид в клетках располагается ассимитрично, на поверхности плазматической мембраны располагается в основном фосфотидолхолины, а внутри фосфотидилколамины – серин.Белки в клеточных мембранах делятся на поверхностные белки и интергральные. Интегральные белки обычно тоже находятся ассимитрично. Толщину мембраны пронизывает гидрофобные обычно уложены в альфа-спирали: С-конец находится на внутренней поверхности, N-конец на внешней поверхности. Очень часто N-концевой фрагмент присоединяет углеводы играют рецепторную функцию. Гидрофобные части белка связываются с гидрофобными участками липидов, а гидрофильные с гидрофильными участками. Физико-химические свойства мембран. Эти свойства определяются химическим составом мембран и температура окружающей среды. Жесткость мембранам придают хлестерин и насыщенные жирные кислоты. Ненасыщенные жирные кислоты придают тянучесть липидам клеточной мембраны. При низкой температуре фосфолипиды достаточно жестко закреплены в составе мембраны, при повышении температуры возможно перемещение липидов. При температуре тела жиры находятся в жидком состоянии.Функции клеточной мембраны.Разделительная – мембраны придают форму клеткам, формируют внутренние отсеки, взаимодействуют с структурой цитоскелета.Коммуникативная – мембраны обеспечивают межклеточные контакты с помощью рецепторов.Метаболическая – в клеточных мембранах встроены мембранные ферменты.Транспортная – через мембрану осуществляется транспорт веществ.Рецепторные – взаимодействие с веществамиТранспорт веществ через клеточные мембраны.Пассивный транспорт веществ который осуществляется по градиенту концентрации через соответствующие клеточные каналыАктивный – при участии белковых переносчиковОблигатный – в котором участвует особые дополнительные транспортные белки - однонаправленный перемещение двух веществ- в различных направлениях4. Транспорт макромолекул осуществляется путем эндоцитоза или экзоцитоза. Функции жиров: Энергетическая: снабжает организм энергией, калорийная ценность жиров выше, чем у углеводов и белков. 1г жира=9 ккал. Энергетическую роль выполняют резервные жирыПластическая: Регуляторная:а) липиды определяют проницаемость клеточных мембран, регулируют активность мембранных ферментовб) из липидов синтезируются особые тканевые гормоны эйкозаноидыЗащитная: липиды создают механическую защиту внутренних органов от повреждений и травмТерморегуляторная: липиды подкожной клетчатки снижают теплоотдачу организмаУчаствуют в проведении нервных импульсов, формируют миелиновые оболочки нервных пучков, стволов => роль электроизолятовЛипиды растворяют жирорастворимые витаминыЖиры являются важными источниками эндогенной воды
БИЛЕТ 30 Na – общее содержание 100 г; 50% внеклеточный натрий, 45% в костях, 5% внутриклеточный. Содержание натрия: плазме крови 130-150 ммль/л, клетках крови 4-10 ммоль/л. Суточная потребность в натрии 4-6 г. К – общее содержание в организме 160 г. 90% внутриклеточный калий, 10% внеклеточный. В плазме 4,5 ммоль/л, Внутри клетки 110 ммоль/лСуточная потребность – 2-4 г/сут. Биологическая роль: Определяют осмотическое давлениеОпределяют распределение водыСоздают артериальное давлениеУчаствуют (Na) в всасывании АК, моносахаров.Калий необходим для биосинтетических процессов.Всасывания Nа и К осуществляется в желудке и в кишечнике. Натрий может незначительно депонироваться в печени. Выводятся натрий и калий в основном через почки, потовые железы и через кишечник.В перераспределении натрия и калия между клетками и внеклеточной жидкостью участвует натрий-калиявая АТФ-аза мембранный фермент который за счет энергии АТФ перемещает ионы натрия и калия против градиента концентрации. Этот перепад концентрации натрия и калия обеспечивает процесс возбуждения ткани. Регуляция водно-солевого обмена Альдостерон – способствует задержке натрия в организме и потере ионов К через почки. Считается что этот гормон способствует синтезу белков определяющих обратную реабсорбцию натрия и синтезу белков натриевых каналов, в почках гормон активирует цикл Кребса, синтез АТФ необходимый для процессов реабсорбции. Активирует синтез транспортеров калия. Функция и АДГ и альдостерона тесно взаимосвязана с особой ренин-ангиотенсиновой системой крови. Ренин-ангиотензивная система крови. При уменьшении кровотока через почки (бывает при уменьшении воды в организме) в почках вырабатывается протеолитический фермент ренин он переводит ангиотензиноген (α2-глобулин) в ангиотензин 1 (пептид в 10 АК)→ангиотензин 2 (8 АК) под воздействием фермента ангиотезин превращающий фермент (АПФ – пептидаза) АГТ2- суживает сосуды, стимулирует выработку АДГ и альдостерона, которые и увеличивают объем жидкости.Противодействующим фактором являтся натрий-уретический пептид (циклический, 28 АК с дисульфидными мостиками. В основном вырабатывается в предсердиях в ответ на увеличение объема воды в организме и на растяжения предсердий. НУП – способствует выведению Nа и воды из организма. Роль почек в водно-солевом обмене. В почках происходит фильтрация, реабсорбции, секреция натрия, калия. На почке оказывает регулирующее влияние АЛЬД, АДГ, в почках вырабатывается ренин, осуществляется выделение протонов и тем самым регулируется рН. Са – общее содержание 2% от массы тела (≈1,5 кг). 99% в костях, 1% внеклеточный. Содержание Са в плазме 2,3-2,8 ммоль/л, 50% этого Са ионизированы (соли), а другие 50% связаны с белками. Функции Са: Пластический материалУчаствует в мышечном сокращенииУчаствует в свертывании кровиРегулятор активности многих ферментов (выполняет роль вторичного посредника.Суточная потребность: 1,5 г. Всасывание Са в ЖКТ лимитировано. Всасывается примерно 50% Са пищевых продуктов при участии кальций-связывающего белка. Р – 1% от массы тела (≈700 г). 90% в костях, 10% внутриклеточный фосфор. В крови 1-2 ммоль/л Роль Р:Пластическая функцияВходит в состав макроэргов (АТФ)Компонент нуклеиновых кислот, липопротеидов, нуклеатизов, солейВходит в состав фосфатного буфера – регуляция рН.Регулятиор активности многих ферментов (фосфолирирование, дефосфолирирования) Суточная потербность: 1,5 г В ЖКТ всасывается при участии щелочной фосфатазы. Выводятся Са и Р в основном через почки незначительно через кишечник.Общее содержание у взрослого 5 г преобладает гемовое железо: гемоглобин, миоглобин, цитохромы. Внеклеточное железо – трансферрин. В плазме: 16-19 мкмоль/л, в эритроцитах 19 ммоль/л. Суточный обмен: взрослый 20-25 мг. 90% этого количества приходится на эндогенное железо освобождающееся при распаде эритроцитов, 10% экзогенное.Функции железа:Обязательный компонент О-В процессов в организмеПеренос кислородаАнтиоксидантная функцияСтимулирует иммунные реакции в организме Всасывание происходит в кишечникеСчитается что всасывается 1/10 часть железа пищевых продуктов. В пищевых продуктах железо 3-х валентное в кислой среде желудка переходит в Fе2+, во всасывании железа участвует белок апоферритин он связывает железо и остается в слизистой кишечника создавая депо. Синтез апоферитина уменьшается при недостатке железа в организме. №2Это процесс гидролитического расщепления поли- и олигосахаридов. Переваривание начинается в ротовой полости. В слюне содержится фермент, расщепляющий крахмал (a-амилаза), расщепляет a-1,4-гликозидные связи внутри молекулы (но не концевые), при этом образуются декстрины, небольшое количество мальтозы и изомальтозы. В желудке углеводы не перевариваются, т.к. рН – кислая. Полное переваривание происходит в тонком кишечнике. Поджелудочная железа вырабатывает a, b - амилазы и выделяет их в просвет кишечника. Клетки слизистой кишечника тоже продуцируют ряд ферментов, переваривающих олигосахара. Одна группа ферментов расщепляет a-гликозидные связи (мальтаза: a-1,4- гликозидные связи, изомальтаза-a-1,6; сахараза- сахарозу). Вторая группа расщепляет b-гликозидные связи (лактаза, b-галактозидаза, гетерогалактозидаза – расщепляет смешанные олигосахариды). Под действием указанных ферментов происходит расщепление до моносахаров. Переваривание чаще идёт пристеночное (у поверхности клеток). Не перевариваются: клетчатка (нет фермента), но она нужна для перистальтики кишечника, пектозаны (полисахара, состоящие из пентоз). При переваривании больше всего образуется глюкозы. Всасывание осуществляется с помощью особых переносчиков, при этом характерна специфичность, затрачивается энергия (повышено потребление кислорода, расходуется АТФ). Глюкоза и галактоза могут всасываться за счёт активного транспорта, за счёт грАДНента концентрации ионов Nа+, создаваемого Nа, К – АТФазой. Это обеспечивает их всасывание даже при низкой концентрации в кишечнике. С током крови по воротной вене глюкоза доставляется в печень, а частично с током крови доставляется во все органы и ткани. поступления и утилизации глюкозы в клетке. Глюкоза может поступить из ЖКТ с током крови, образоваться при распаде гликогена, может быть синтезирована из неуглеводных компонентов – глюконеогенез. Утилизация глюкозы: Синтез гликогена, синтез жиров, синтез аминокислот, синтез других углеводов,полностью может расщепляться до СО2 и Н2О с образованием энергии.
|
|||||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 211; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.115 (0.012 с.) |

 1) АК + АТФ аминоацил – АМФ + ПФ
1) АК + АТФ аминоацил – АМФ + ПФ 2) аминоацил-АМФ + тРНК аминоацил-тРНК
2) аминоацил-АМФ + тРНК аминоацил-тРНК





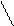 это плечо необходимо это плечо участвует в связывании аминоацил-
это плечо необходимо это плечо участвует в связывании аминоацил-