12. Сопряжение окисления и фосфорилирования. Точки сопряжения. Укороченный вариант цепи биологического окисления
Сопряженное окисление - окисление, при котором энергия окислительной реакции используется для синтеза АТФ. Поэтому данный вид окисления называют окислением, сопряженным с фосфорилированием АДФ. Он может осуществляться двумя способами.
Если при окислении субстрата образуется макроэргическое соединение, энергия которого используется для синтеза АТФ, то такой вид биологического окисления называют субстратным фосфорилированием или фосфорилированием на уровне субстрата или окислением, сопряженным с фосфорилированием АДФ на уровне субстрата. Примером таких реакций являются 2 реакции гликолиза: превращение 1,3-дифосфоглицериновой кислоты в 3-фосфоглицериновую кислоту и фосфоенолпирувата (ФЕП) в пируват, а также реакция цикла Кребса – гидролиз сукцинил-КоА до сукцината. Эти реакции протекают сопряжено с синтезом АТФ.
Если процессы окисления, протекающие в дыхательной цепи ферментов на внутренней мембране митохондрий, где происходит перенос протонов и электронов от окисляемого субстрата к кислороду, сопряжены с синтезом АТФ, то такой вид биологического окисления называют окислительным фосфорилированием или фосфорилированием на уровне электронно-транспортной цепи.
Схема классификации биологического окисления
Биологическое окисление
Свободное окисление Сопряженное окисление
Субстратное Окислительное
фосфорилирование фосфорилирование
В окислительном фосфорилировании используются реакции дегидрогенирования окисляемого субстрата с последующим переносом атомов водорода (протонов и электронов) на кислород при участии оксидоредуктаз. Перенос водорода на кислород происходит через ряд окислительно-восстановительных систем, которые располагаются в строгой последовательности - в соответствии со значением их потенциала. Такая последовательность реакций, связанных с переносом водорода на кислород при участии специфических переносчиков электронов, называется дыхательной (или электронно-транспортной) цепью. У животных и человека она составлена из четырех основных типов переносчиков, каждый из которых способен претерпевать обратимое окисление и восстановление в результате потери и присоединения электронов при взаимодействии с другим переносчиком
Рис. 2. Взаимное расположение компонентов дыхательной цепи с указанием мест фосфорилирования и специфических ингибиторов.
 13. Объясните механизм окислительного фосфорилирования (хемиосмотическая гипотеза) и формирование электрохимического потенциала при переносе электронов по дыхательной цепи.
13. Объясните механизм окислительного фосфорилирования (хемиосмотическая гипотеза) и формирование электрохимического потенциала при переносе электронов по дыхательной цепи. 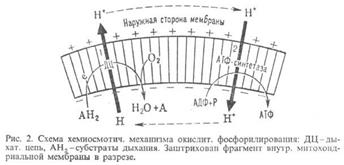
Механизм окислительного фосфорилирования можно представить схемой: Перенос электронов (дыхание)  А ~ В
А ~ В  АТФ А ~ В-высокоэнергетич.интермедиат Предполагалось, что А ~ В - хим. соед. с макроэргич. связью, напр. фосфорилир. фермент дыхат. цепи (хим. гипотеза сопряжения), или напряженная конформация к.-л. белка, участвующего в окислительном фосфорилировании (конформац. гипотеза сопряжения). Однако эти гипотезы не получили эксперим. подтверждения. Наиб. признанием пользуется хемиосмотич. концепция сопряжения, предложенная в 1961 П. Митчеллом (за развитие этой концепции в 1979 ему присуждена Нобелевская премия). Согласно этой теории, своб. энергия транспорта электронов в дыхат. цепи затрачивается на перенос из митохондрий через митохондриальную мембрану на ее наружную сторону ионов Н+ (рис. 2, процесс 1). В результатена мембране возникает разность электрич. потенциалов
АТФ А ~ В-высокоэнергетич.интермедиат Предполагалось, что А ~ В - хим. соед. с макроэргич. связью, напр. фосфорилир. фермент дыхат. цепи (хим. гипотеза сопряжения), или напряженная конформация к.-л. белка, участвующего в окислительном фосфорилировании (конформац. гипотеза сопряжения). Однако эти гипотезы не получили эксперим. подтверждения. Наиб. признанием пользуется хемиосмотич. концепция сопряжения, предложенная в 1961 П. Митчеллом (за развитие этой концепции в 1979 ему присуждена Нобелевская премия). Согласно этой теории, своб. энергия транспорта электронов в дыхат. цепи затрачивается на перенос из митохондрий через митохондриальную мембрану на ее наружную сторону ионов Н+ (рис. 2, процесс 1). В результатена мембране возникает разность электрич. потенциалов  и разность хим. активностей ионов Н+
и разность хим. активностей ионов Н+ 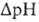 (внутри митохондрий рН выше, чем снаружи). В сумме эти компоненты дают трансмембранную разность электрохим. потенциалов ионов водорода
(внутри митохондрий рН выше, чем снаружи). В сумме эти компоненты дают трансмембранную разность электрохим. потенциалов ионов водорода  между матриксоммитохондрий и внеш. водной фазой, разделенными мембраной:
между матриксоммитохондрий и внеш. водной фазой, разделенными мембраной:

где R-универсальная газовая постоянная, T-абс. т-ра, F- число Фарадея. Величина  обычно составляет ок. 0,25 В, причем осн. часть (0,15-0,20 В) представлена электрич. составляющей
обычно составляет ок. 0,25 В, причем осн. часть (0,15-0,20 В) представлена электрич. составляющей  . Энергия
. Энергия  , выделяющаяся при движении протонов внутрь митохондрий по электрич. полю в сторону меньшей их концентрации (рис. 2, процесс 2), используется АТФ-синтетазой для синтеза АТФ. Т. обр., схему окислительногофосфорилирования, согласно этой концепции, можно представить в след. виде:
, выделяющаяся при движении протонов внутрь митохондрий по электрич. полю в сторону меньшей их концентрации (рис. 2, процесс 2), используется АТФ-синтетазой для синтеза АТФ. Т. обр., схему окислительногофосфорилирования, согласно этой концепции, можно представить в след. виде:
Перенос электронов (дыхание)  АТФ
АТФ
Сопряжение окисления и фосфорилирования через  позволяет объяснить, почему окислительное фосфорилирование, в отличие от гликолитич. ("субстратного") фосфорилирования, протекающего в р-ре, возможно лишь в замкнутых мембранных структурах, а также почему все воздействия, снижающие электрич. сопротивление и увеличивающие протонную проводимость мембраны, подавляют ("разобщают") окислительноефосфорилирование. Энергия
позволяет объяснить, почему окислительное фосфорилирование, в отличие от гликолитич. ("субстратного") фосфорилирования, протекающего в р-ре, возможно лишь в замкнутых мембранных структурах, а также почему все воздействия, снижающие электрич. сопротивление и увеличивающие протонную проводимость мембраны, подавляют ("разобщают") окислительноефосфорилирование. Энергия  , помимо синтеза АТФ, может непосредственно использоваться клеткой для др. целей - транспортаметаболитов, движения (у бактерий), восстановления нико-тинамидных коферментов и др.
, помимо синтеза АТФ, может непосредственно использоваться клеткой для др. целей - транспортаметаболитов, движения (у бактерий), восстановления нико-тинамидных коферментов и др.
В дыхат. цепи имеется неск. участков, к-рые характеризуются значит. перепадом окислит.-восстановит. потенциала 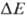 и сопряжены с запасанием энергии (генерацией
и сопряжены с запасанием энергии (генерацией  ). Таких участков, наз. пунктами или точками сопряжения, обычно три: НАДН: убихинон-редуктазное звено (
). Таких участков, наз. пунктами или точками сопряжения, обычно три: НАДН: убихинон-редуктазное звено ( 0,35-0,4 В), убихинол: цитохром-c-редуктазное звено (
0,35-0,4 В), убихинол: цитохром-c-редуктазное звено ( ~ ~ 0,25 В) и цитохром-с-оксидазный комплекс (
~ ~ 0,25 В) и цитохром-с-оксидазный комплекс ( ~ 0,6 В)-пункты сопряжения 1, 2 и 3 соотв. (рис. 3). Каждый из пунктов сопряжения дыхат. цепи м.б. выделен из мембраны в виде индивидуального ферментного комплекса, обладающего окислит.-восстановит. активностью. Такой комплекс, встроенный в фосфолипидную мембрану, способен функционировать как протонный насос.
~ 0,6 В)-пункты сопряжения 1, 2 и 3 соотв. (рис. 3). Каждый из пунктов сопряжения дыхат. цепи м.б. выделен из мембраны в виде индивидуального ферментного комплекса, обладающего окислит.-восстановит. активностью. Такой комплекс, встроенный в фосфолипидную мембрану, способен функционировать как протонный насос.

Обычно для характеристики эффективности окислительного фосфорилирования используют величины Н+/2е или q/2e, указывающие сколькопротонов (либо электрич. зарядов) переносится через мембрану при транспорте пары электронов через данный участок дыхат. цепи, а также отношение Н+/АТФ, показывающее, сколько протонов нужно перенести снаружи внутрь митохондрий через АТФ-синтетазу для синтеза 1 молекулыАТФ. Величина q/2e составляет для пунктов сопряжения 1, 2 и 3 соотв. 3-4, 2 и 4. Величина Н+/АТФ при синтезе АТФ внутри митохондрий равна 2; однако еще один Н+ может тратиться на вынос синтезированного АТФ4- из матрикса в цитоплазму переносчиком адениновых нуклеотидов в обмен на АДФ -3. Поэтому кажущаяся величина Н+ / АТФнаружн равна 3.





