Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
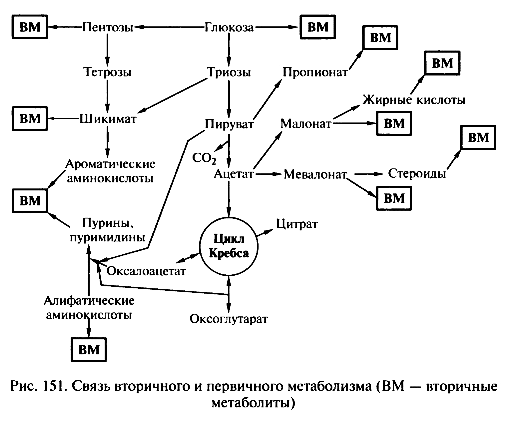
Синтез основных биополимеров и важнейших компонентов клеток
Белки синтезируются из двадцати аминокислот, предшественниками которых являются различные интермедиаты катаболизма. Все аминокислоты делятся на группы в соответствии со своим биосинтетическим происхождением. Синтез аминокислот группы · глутаминовой к-ты (глутаминовая к- та, глутамин, аргинин, пролин) берет начало от альфа-кетоглутарата, интермедиата цикла Кребса. · Оксалоацетат дает начало цепи реакций, приводящих к образованию аспарагиновой кислоты, аспарагина, метионина, треонина, изолейцина и лизина (группа аспарагиновой кислоты). · Синтезы группы ароматических аминокслот (триптофана, фенилаланина и тирозина) начинаются с конденсации ФЕП из гликолитического пути и эритрозо-4-фосфата. · 3-ФГК и пируват дают начало реакциям, приводящим к синтезу аминокислот группы серина (серин, глицин, цистеин) и группы пировиноградной кислоты (аланин, валин, лейцин) соответственно. · Биосинтез гистидина тесно связан с путями образования пуринов. Два атома углерода пятичленного имидазольного кольца и три углерода боковой цепи происходят из фосфорибозилпирофосфата. Фрагмент C—N этого кольца образуется из пуринового ядра АТФ, а другой атом азота — из глутамина.
· Для биосинтеза белков требуется присутствие не только ферментов и мономеров, но и матрицы (молекулы иРНК), задающей последовательность присоединения аминокислот к растущей цепи, а также специфического переносчика для активирования мономера и отбора его в соответствии с заданным кодом (тРНК).
Липиды. Группа соединений, разнообразных по химическому составу, но нерастворимых в воде. Их можно условно разделить на вещества: · содержащие жирные кислоты, связанные эфирной связью (нейтральные жиры, фосфолипиды, гликолипиды, липополисахариды, полиалканоаты и т.д.), · и вещества, содержащие изопреновые фрагменты (полиизопрены, каротиноиды, стеролы, хлорофиллы, хиноны и т.д.).
Наращивание углеродной цепи образуемой жирной кислоты происходит на ацилпереносящем белке (АПБ). Синтез жирных кислот с четным и нечетным числом атомов отличается только первой реакцией, где происходит конденсация пропионил-АПБ с амонил-АПБ.
В синтезе фосфолипидов принимает участие интермедиат гликолиза диоксиацетонфосфат, который восстанавливается до 3- фосфоглицерола и присоединяет два остатка жирных кислот, связанных с АПБ.
Первые стадии образования изопреноидных липидов связаны с последовательной конденсацией трех молекул ацетил-КоА в разных положениях и перегруппировкой полученного соединения. В результате синтезируется разветвленная мевалоновая кислота, которая претерпевает два последовательных фосфорилирования и декарбоксилирование с образованием активированного С5-соединения, являющегося предшественником сложных изопреноидных веществ. Сложные липиды могут строиться из разнообразных предшественников – жирных кислот, многоатомных спиртов, изопренов, а также моносахаров, аминокислот и аминов. Высшие жирные кислоты синтезируются из активированной уксусной кислоты, предшественником других липидных макромолекул может быть изопреновый фрагмент.
Нуклеиновые кислоты. Синтезируются и з пурин - и пиримидиннуклеозидтрифосфатов(AТФ, ГТФ, ЦТФ, ТТФ, УТФ), которые имеют в целом сходное строение. В них пуриновое или пиримидиновое основание соединено с пентозой через атом азота (нукоеозид), а фосфатные группы находятся в 5’-положении (нуклеотид). Дезоксирибонуклеотиды обр. из рибонуклеотидов путем восстановления. Синтез рибонуклеотидов начинается с образования 5-фосфорибозил-1-пирофосфата. Рибозо-5-фосфат является промежуточным продуктом пентозофосфатного пути. Пуриновые рибонуклеотиды далее синтезируются путем последовательного присоединения амино- и углеродсодержащих групп к ФРПФ с образованием девятичленного пуринового кольца. В случае пиримидиновых рибонуклеотидов снчала происходит конденсация аспарагиновой кислоты и карбамоилфосфата с образованием шестичленного пиримидинового кольца, а затем присоединение рибозофосфатного остатка. Образование нуклеиновых кислот осущ. путем матричного синтеза в процессах репликации и транскрипции.
Порфирины. Важной группой изопреноидных соединений являются порфирины – сложные азотсодержащие вещества, служащие простетическими группами ряда ферментов и хлорофиллов. Конденсация сукцинил-КоА и глицина и ряд по след ующи х модификаций приводя т к обра зованию пятичленного азотсодержащего кольца с тремя заместителями (порфобилиногена), четыре молекулы которого далее объединяются в тетрапиррольное ядро протопорфирина IX. Последующие реакции могут приводить к синтезу гема (при введении в молекулу атома железа) или образованию хлорофилла (при введении в молекулу магния). Различные формы хлорофиллов синтезируются в дальнейшем с помощью реакций периферического метаболизма определенных групп фототрофных микрооорганизмов.
Пептидоглюкан=муреин. Состоит ил линейных молекул гликана, соединенных между собой поперечно пептидами с аминокислотными остатками. Гликан муреина образован n-ацетил-n-глюкозамином и n-ацетилмурамовой кислотой через бета-1-4-связи.
Синтез: (n-ацетил-n-глюкозамином + уридиндифосфат) + ФЕП = n-ацетилмурамовая кислота Потом с помощьэ ферментов и затратой АТФ формируются мостики при наличии ионов Mg. Далее в ЦПМ происходит реакция трансгликозилирования при участии фосфолипида-переносчика.
Вторичные метаболиты. Вторичные метаболиты очень разнообразны. Каждый вид второчного метаболита производит небольшой круг видов. Вторичные метаболиты -- вещества микробного (или растительного) происхождения, не существенные для роста и репродукции организма. Вообще, строгого определения нет. ВМ синтезируются в конце экспоненциальной или в течение стационарной фазы роста, а их формирование зависит не только от штамма микроорганизма, но и от условий роста. ВМ относятся к различным классам химических соединений:аминоциклитолы, кумарины, эпоксиды, полиены, пирролыв, терпеноиды, тетрациклины, нерибосомальные пептиды и др. Важной группой ВМ являются антибиотики: например, эрготоксин спорыньи, поликетиды (которые не только антибиотики, но и иммунодепрессанты широкого спектра), изопреноиды (каротиноиды, стероиды, гиббереллины, фитоалексины, являющиеся фитогормонами, антираковые антибиотики). к ВМ также относятся полигидроксиалканоаты, являющиеся запасающими веществами. Все вторичные метаболиты синтезируются из первичных метаболитов. Способы получения ВМ: 1) Преобразование первичного метаболита в специфические предшественники для ВМ 2) реакция модификации или актвации, уводящие предшественник на путь вторичного меиаболизма 3) полимеризация и конденсация 4) поздние реакции модификации
Первичные метаболиты синтезируются в трофофазе, когда происходит рост и размножение колонии, а синтез ВМ -- в идиофазе (замежление роста, остановка роста и начала синтеза продуктов)
Образование антибиотиков микроорганизмами. Вторичные метаболиты. Это вещества микробного (или растительного) происхождения, не существенные для роста и репродукции образующего их организма. Каждый вторичный метаболит производится относительно ограниченным числом видов. Эти соединения синтезируются в конце экспоненциальной или в течение стационарной фаз роста, и их формирование взначительной степени зависит от условий роста, особенно состава питательной среды. Многие вторичные метаболиты имеют химическую структуру, необычную для биологической материи, однако их образование берет начало от интермедиатов первичного метаболизма. В отличие от синтеза первичного метаболита, который происходит одновременно с ростом и размножением культуры, для продуцента вторичных метаболитов принято говорить о трофофазе (когда культура растет и размножается) и идиофазе (когда рост замедляется или останавливается и начинается синтез продукта). Механизмы переключения путей метаболизма с первичного на вторичный не ясны.
К вторичным метаболитам причисляют антибиотики, токсины, иммунодепрессанты и стимуляторы, а также некоторые запасные вещества (поли-β-алканоаты). Эти соединения относятся к разнообразным классам органических веществ (аминоциклитолы, кумарины, эпоксиды, нерибосомальные пептиды, полиены, пирролы, терпеноиды, тетрациклины, поликетиды, изопреноиды, стероиды, гиббереллины, фитоалексины и т.д.). О физиологической роли вторичных метаболитов в жизни собственного продуцента достоверно известно очень мало. Неизвестно, насколько распространен вторичный метаболизм в природе. Само понятие “вторичный метаболит” достаточно расплывчатое и многие исследователи его не признают.
Антибиотики - вторичные метаболиты. Нахрен бактериям это сдалось: Антагонизм – подавление роста конкурентов. Среди бактерий наиболее часто явление антагонизма встречается у Bacillus. Из культуры неспороносной бактерии антибактериальное вещество пиоцианаза было выделено Р. Эммерихом и О. Лоу в конце прошлого века. Позже были обнаружены антибиотические вещества в культурах Bacterium prodigiosum, молочнокислых стрептококков, микрококков, азотобактера и др. Примеры: Bacillus brevis (грамицидин и тироцидин), Bacillus brevis var. G.-B. (грамицидин С), Bacillus subtilis и Вас. mesentericus (субтилин, бацитрацин), Bacillus polymyxa (полимиксин), Pseudomonas pyocyanea (пиоцианин), Streptococcus lactis (низин). Самой богатой антагонистами группой почвенных микроорганизмов оказалась группа лучистых грибков, актиномицетов, а среди них — представители рода Actinomyces. Подавляющее большинство применяющихся антибиотиков, получено именно из этой группы.
|
|||||||
|
|
Последнее изменение этой страницы: 2016-12-30; просмотров: 795; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.112.210 (0.012 с.) |