Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Схема синтеза белка в эукариотной клетке.
1. Транскрипция Отдельные участки двутяжевой ДНК, называемые генами, являются матрицами для синтеза на них однотяжевых цепей РНК. Синтезированные цепи РНК комплементарны одной из цепей ДНК и, таким образом, точно воспроизводят дезоксирибонуклеотидную последовательность другой цепи ДНК в своей рибонуклеотидной последовательности. 2. Процессинг и транспорт РНК РНК в течение синтеза и после него, особенно в эукариотических клетках, может подвергаться ряду дополнительных изменений (добавление концевых групп, модификации нуклеотидов, вырезание определенных кусков нуклеотидной последовательности и др.). Получающаяся информационная, или мессенджер, РНК (мРНК) поступает далее к рибосомам (у эукариот транспортируется из ядра в цитоплазму) в качестве программы, определяющей аминокислотную последовательность в синтезируемом белке. Активация и акцептирование аминокислот. Исходным материалом, из которого строится белок, являются аминокислоты, однако свободные аминокислоты клетки не могут быть непосредственно использованы рибосомой. Каждая аминокислота сначала активируется с помощью АТФ, а затем присоединяется к специальной молекуле РНК, называемой трансферной РНК (тРНК), вне рибосомы. Получающаяся аминоацил-тРНК поступает в рибосому в качестве субстрата для синтеза белка. 3. Трансляция Поток информации в виде мРНК и поток материала в виде аминоацил-тРНК поступают в рибосомы, которые являются молекулярными машинами, осуществляющими перевод, или трансляцию, генетической информации с языка нуклеотидной последовательности мРНК на язык аминокислотной последовательности синтезируемой полипептидной цепи белка. Каждая рибосома последовательно сканирует цепь мРНК (движется вдоль нее от одного конца к другому) и соответственно выбирает из среды те аминоацил- тРНК, которые соответствуют (комплементарны) триплетным комбинациям нуклеотидов, находящимся в данный момент в рибосоме. Таким образом, движение рибосомы вдоль мРНК задает строгий временной порядок вхождения в рибосому разных аминоацил-тРНК в соответствии с порядком расположения кодирующих нуклеотидных комбинаций (кодонов) вдоль мРНК. Аминокислотный остаток выбранной аминоацил-тРНК каждый раз ковалентно присоединяется рибосомой к растущей полипептидной цепи, а деацилированная тРНК освобождается из рибосомы в раствор. Так последовательно остаток за остатком строится полипептидная цепь.
4. Формирование функционального белка По мере синтеза полипептидная цепь частично высовывается из рибосомы и начинает сворачиваться в глобулу (котрансляционный фолдинг), а по завершении синтеза, то есть по прочтении всей мРНК, она освобождается из рибосомы и окончательно сворачивается (посттрансляционный фолдинг). Синтезируемый белок может транспортироваться через клеточные мембраны, что характерно для белков, производимых клеткой для общих нужд организма или клеточной популяции (секретируемые белки). Сворачивание белка и транспорт белка через мембраны может сопровождаться различными ковалентными модификациями с помощью ферментов (процессинг белка) Итак, процесс создания химической структуры белка (синтез полипептидной цепи) и в значительной мере ее физическое сворачивание в функционально активную белковую глобулу осуществляются рибосомой. Количество рибосом в клетке сильно варьирует - от тысяч до десятков тысяч на клетку - в зависимости от интенсивности белкового синтеза в данном типе клеток. Каждая рибосома полностью прочитывает одну молекулу мРНК и в соответствии с ее программой синтезирует одну молекулу белка, после чего может быть запрограммирована другой молекулой мРНК и произвести другую молекулу белка и т.д. Обычно одна молекула мРНК читается сразу несколькими рибосомами, двигающимися вдоль мРНК друг за другом и, таким образом, независимо синтезирующими идентичные молекулы белка, но с соответствующим отставанием. Такой динамический комплекс одной мРНК с несколькими рибосомами называется полирибосомой. Химически рибосома есть рибонуклеопротеид: она состоит из специальной рибосомной РНК и специальных рибосомных белков, находящихся в комплексе друг с другом. Физически рибосома представляет собой компактную частицу специфической формы, лишенную внутренней и внешней симметрии, грубо аппроксимируемую сферой с диаметром около 30 нм. Функционально это молекулярная машина, протягивающая вдоль себя цепь мРНК, считывающая закодированную в мРНК генетическую информацию и параллельно, в соответствии с кодом, синтезирующая полипептидную цепь белка из поступающих в нее аминокислотных остатков. В процессе работы рибосома потребляет энергию гидролиза гуанозинтрифосфата (ГТФ). Очевидно, что детальное знание структуры рибосомы является необходимой базой для понимания механизмов работы этой молекулярной машины. В настоящее время полная структура рибосомы на молекулярном уровне еще неизвестна, хотя известно много деталей ее строения. В этой статье делается попытка обобщить многочисленные разрозненные сведения о структуре рибосом и сформулировать основные принципы, лежащие в основе ее молекулярной организации.
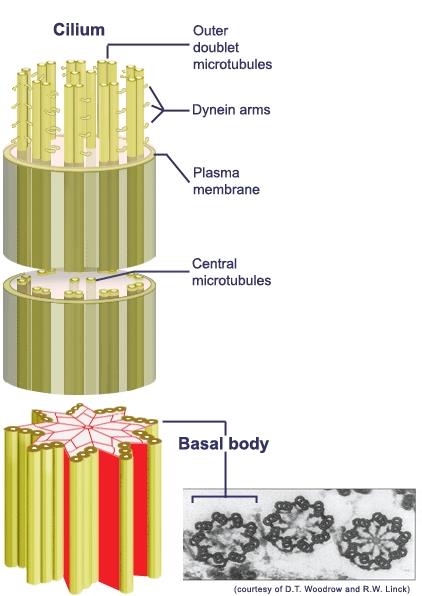
Микротрубочками. Во многих клетках обнаружены органоиды, названные микротрубочками. Само их название говорит об их форме — это трубочки с каналом внутри. Внешний их диаметр порядка 250Å. Иногда это двойные трубочки — две одиночные, лежащие бок о бок друг с другом и имеющие общую стенку, которая разделяет их полости. Стенки микротрубочек построены из белковых молекул. Микротрубочки связаны с сократительной (двигательной) активностью цитоплазмы и ее образований. Из них, как из строительных деталей, построены сократительные структуры жгутика — органоида, при помощи которого перемещаются некоторые одноклеточные и колониальные водоросли, а также клетки, служащие для размножения многих низших растений. Микротрубочки формируют центральную структуру ресничек и жгутиков — аксонему. Типичная аксонема содержит 9 пар объединённых микротрубочек по периферии и две полных микротрубочки в центре
Из микротрубочек состоят также центриоли и веретено деления, обеспечивающее расхождение хромосом к полюсам клетки при митозе и мейозе. В период деления микротрубочки собираются в группы и образуют эти нити. По окончании деления нити вновь распадаются на отдельные микротрубочки. В клетках или их частях, которые лишены плотной оболочки, микротрубочки выполняют опорную функцию, составляя внутренний скелет клетки. Микротрубочки в клетке используются в качестве «рельсов» для транспортировки частиц. По их поверхности могут перемещаться мембранные пузырьки и митохондрии. Транспортировку по микротрубочкам осуществляют белки, называемые моторными. Это высокомолекулярные соединения, состоящие из двух тяжёлых (массой около 300 кДа) и нескольких лёгких цепей. В тяжёлых цепях выделяют головной и хвостовой домены. Два головных домена связываются с микротрубочками и являются собственно двигателями, а хвостовые — связываются с органеллами и другими внутриклеточными образованиями, подлежащими транспортировке. Выделяют два вида моторных белков: цитоплазматические динеины; кинезины. Динеины перемещают груз только от плюс-конца к минус-концу микротрубочки, то есть из периферийных областей клетки к центросоме. Кинезины, напротив, перемещаются к плюс-концу, то есть к клеточной периферии. Перемещение осуществляется за счёт энергии АТФ. Головные домены моторных белков для этого содержат АТФ-связывающие участки. Микротрубочки участвуют в поддержании формы клетки и расположения органоидов (в частности, аппарата Гольджи) в цитоплазме клеток.
Пластиды Пластиды — органоиды, присущие только растительным клеткам. Обычно это крупные тельца, хорошо видимые под световым микроскопом.
Хлоропласт из клетки листа огурца. Электронная микрофотография (увел. х48 000): о — оболочка хлоропласта; л — межгранные ламеллы; гр — граны; с — строма; кз — крахмальные зерна; ог — осмиофильные гранулы (капли жироподобных веществ); м — митохондрия.
Фрагмент хлоропласта из пареихимной клетки листа кукурузы. Электронная микрофотография (увел. х100 000): о — оболочка хлоропласта; л — межгранные ламеллы; г — граны; с — строма; кз — крахмальное зерно; oг — осмиофильные гранулы.
Различают 3 типа пластид: Лейкопласты — неокрашенные пластиды, как правило выполняют запасающую функцию. В лейкопластах клубней картофеля накапливается крахмал. Лейкопласты высших растений могут превращаться в хлоропласты или хромопласты. Хромопласты — пластиды, окрашенные в жёлтый, красный или оранжевый цвет. Окраска хромопластов связана с накоплением в них каротиноидов. Хромопласты определяют окраску осенних листьев, лепестков цветов, корнеплодов, созревших плодов. Хлоропласты — пластиды, несущие фотосинтезирующие пигменты — хлорофиллы. Имеют зелёную окраску у высших растений, харовых и зелёных водорослей. Набор пигментов, участвующих в фотосинтезе (и, соответственно, определяющих окраску хлоропласта) различен у представителей разных таксономических отделов. Хлоропласты имеют сложную внутреннюю структуру. Пластиды каждого типа имеют свое строение и несут свои, им присущие функции. Однако возможны переходы пластид из одного типа в другой. Так, позеленение клубней картофеля вызывается перестройкой их лейкопластов в хлоропласты. В корнеплоде моркови лейкопласты переходят в хромопласты. Пластиды всех трех типов образуются из пропластид. Пропластиды — бесцветные тельца, похожие на митохондрии, но несколько крупнее их. В больших количествах они встречаются в меристематических клетках. Лейкопласты находятся в клетках неокрашенных частей растений (плодов, семян, корней, эпидермиса листьев). Форма их неопределенна. Чаще всего встречаются лейкопласты, в которых откладывается крахмал (он образуется из сахаров). Есть лейкопласты, запасающие белки. Наименее распространены лейкопласты, заполненные жиром; они образуются при старении хлоропластов. Существенных различий между лейкопластами и пропластидами нет.
Хлоропласты — пластиды высших растений, в которых идет процесс фотосинтеза, т. е. использование энергии световых лучей для образования из неорганических веществ (углекислого газа и воды) органических веществ с одновременным выделением в атмосферу кислорода. Хлоропласты имеют форму двояковыпуклой линзы, размер их около 4-6 мкм. Находятся они в паренхимных клетках листьев и других зеленых частей высших растений. Число их в клетке варьирует в пределах 25-50.
Строение хлоропласта.
Снаружи хлоропласт покрыт оболочкой, состоящей из двух липопротеиновых мембран. Под ней, в основном веществе (строме), упорядоченно расположены многочисленные образования — ламеллы. Они образуют плоские мешочки, которые лежат друг на друге правильными стопками. Эти стопки, напоминающие монеты, сложенные столбиком, называются гранами. Сквозь них проходят более длинные ламеллы, так что все граны хлоропласта связаны в единую систему. В состав мембран, образующих граны, входит зеленый пигмент — хлорофилл. Именно здесь происходят световые реакции фотосинтеза — поглощение хлорофиллом световых лучей и превращение энергии света в энергию возбужденных электронов. Электроны, возбужденные светом, т. е. обладающие избыточной энергией, отдают свою энергию на разложение воды и синтез АТФ. При разложении воды образуются кислород и водород Далее, с помощью ферментов стромы хлоропластов, т. е. вне гран, протекают темновые реакции: водород и энергия, заключенная в АТФ, используются для восстановления атмосферного углекислого газа (CO2) и включения его при этом в состав органических веществ. Первое органическое вещество, образующееся в результате фотосинтеза, подвергается большому числу перестроек и дает начало всему многообразию органических веществ, синтезирующихся в растении и составляющих его тело. Ряд из этих превращений происходит тут же, в строме хлоропласта, где имеются ферменты для образования Сахаров, жиров, а также все необходимое для синтеза белка. Сахара могут затем либо перейти из хлоропласта в другие структуры клетки, а оттуда в другие клетки растения, либо образовать крахмал, зерна которого часто можно видеть в хлоропластах. Жиры тоже откладываются в хлоропластах или в виде капель, или в форме более простых веществ, предшественников жиров, выходят из хлоропласта. Усложнение веществ сопряжено с созданием новых химических связей и обычно требует затрат энергии. Источник ее — все тот же фотосинтез. Значительная доля веществ, образующихся в результате фотосинтеза, вновь распадается в гиалоплазме и митохондриях (в случае полного сгорания — до веществ, которые служат исходным материалом для фотосинтеза, — CO2 и H2O). В результате этого процесса, по своей сути обратного фотосинтезу, энергия, ранее аккумулированная в химических связях разлагаемых веществ, освобождается и — снова через посредство АТФ — тратится на образование новых химических связей синтезируемых молекул. Таким образом, существенная часть продукции фотосинтеза нужна только для того, чтобы связать энергию света и, превратив ее в химическую, использовать для синтеза совсем других веществ. И лишь часть органического вещества, образующегося при фотосинтезе, используется как строительный материал для этих синтезов.
Продукция фотосинтеза (биомасса) колоссальна. За год на земном шаре она составляет около 1010 т. Органические вещества, создаваемые растениями, — это единственный источник жизни не только растений, но и животных, так как последние перерабатывают уже готовые органические вещества, питаясь либо непосредственно растениями, либо другими животными, которые, в свою очередь, питаются растениями. Таким образом, в основе всей современной жизни на Земле лежит фотосинтез. Все превращения веществ и энергии в растениях и животных представляют собой перестройки, перекомбинации и переносы вещества и энергии первичных продуктов фотосинтеза. Фотосинтез важен для всего живого и тем, что одним из его продуктов является свободный кислород, происходящий из молекулы воды и выделяющийся в атмосферу. Полагают, что весь кислород атмосферы образовался благодаря фотосинтезу. Он необходим для дыхания как растениям, так и животным. Хлоропласты способны перемещаться по клетке. На слабом свету они располагаются под той стенкой клетки, которая обращена к свету. При этом они обращаются к свету своей большей поверхностью. Если свет слишком интенсивен, они поворачиваются к нему ребром и; выстраиваются вдоль стенок, параллельных лучам света. При средних освещенностях хлоропласты занимают положение, среднее между двумя крайними. В любом случае достигается один результат: хлоропласты оказываются в наиболее благоприятных для фотосинтеза условиях освещения. Такие перемещения хлоропластов (фототаксис) — это проявление одного из видов раздражимости у растений. Хлоропласты обладают известной автономией в системе клетки. В них имеются собственные рибосомы и набор веществ, определяющих синтез ряда собственных белков хлоропласта. Имеются также ферменты, работа которых приводит к образованию липидов, входящих в состав ламелл, и хлорофилла. Как мы видели, хлоропласт располагает и автономной системой добывания энергии. Благодаря всему этому хлоропласты способны самостоятельно строить собственные структуры. Существует даже взгляд, что хлоропласты (как и митохондрии) произошли от каких-то низших организмов, поселившихся в растительной клетке и сперва вступивших с нею в симбиоз, а затем ставших ее составной частью, органоидом. У низших растений фотосинтез также осуществляется специализированными, хотя и не столь высокоразвитыми, как в хлоропласте, мембранными структурами. У фотосинтезирующих бактерий мембраны, содержащие хлорофилл, образуют сеть, которая пронизывает тело бактерии. У сине-зеленых водорослей фотосинтезирующие мембраны слиты в плоские пузырьки. У зеленых и других водорослей система этих мембран отделена от остальной части клетки покрывающей мембраной и образует специальный органоид — хроматофор. Число хроматофоров в клетке невелико, часто клетка содержит всего лишь один хроматофор. Форма их очень различна у водорослей разных видов. У спирогиры хроматофор имеет вид ленты, спирально вьющейся вдоль стенок клетки; у клостридиума — это ребристые цилиндры; у зигнемы — звездчатые тела.
Хромопласты окрашенные внутриклеточные органеллы растительных клеток, тип пластид. Бывают шарообразными, веретеновидными, серповидными и неправильно-многоугольными. Окраска (оранжевая, жёлтая или буроватая) зависит в основном от присутствия в содержимом пигментов каротиноидов. Обычно образуются из зелёных пластид — хлоропластов вследствие разрушения в них зелёных пигментов — хлорофиллов в процессе созревания плодов некоторых растений (рябины, ландыша, хурмы и др.), а также осеннего пожелтения листьев. При этом происходит распад белково-липидной мембранной системы хлоропластов. Белковый компонент оттекает из пластид, а липидный остаётся внутри. В нём растворяются каротиноиды и окрашивают пластиды в оранжевые и жёлтые тона. В некоторых случаях возникают из бесцветных пластид — лейкопластов (например, в корнеплодах моркови). Их внутренняя мембранная структура гораздо проще, чем у хлоропластов. Гран нет, строма содержит много желтого или оранжевого пигмента. Хромопласты содержатся в клетках лепестков, плодов, корнеплодов. Лейкопласты. Пластиды, не содержащие в строме пигментов и называемые лейкопластами, имеются во многих клетках большинства растений. Так как лейкопласты бесцветны и к тому же преломляют свет почти так же, как протоплазма, не всегда легко обнаружить их присутствие в клетке. По форме лейкопласты обычно почти шаровидны. В тех случаях, когда в их строме находится крахмал или белок, они принимают иные очертания. Сравнительно богаты лейкопластами образовательные ткани, подземные органы, семена. В лейкопластах может образовываться крахмал, отлагающийся в виде зерен в их строме. Во многих случаях крахмал накопляется в лейкопласте в столь большом количестве, что живое тело пластиды (строма) оттесняется на периферию. Его можно не без труда заметить в виде очень тонкой пленки на поверхности крахмального зерна; в этих случаях лейкопласт является крахмалонакопителем в полной мере.
Вакуоль В типичной растительной клетке имеется крупная вакуоль, наполненная жидким содержимым. Часто вакуоль занимает почти весь объем клетки, так что цитоплазма составляет лишь тонкий слой, прилегающий к клеточной оболочке. У молодых клеток бывает несколько мелких вакуолей, которые по мере развития клетки разрастаются и сливаются в одну. Вакуоли - крупные мембранные пузырьки или полости в цитоплазме, заполненные клеточным соком. Вакуоли образуются в клетках растений и грибов из пузыревидных расширений эндоплазматического ретикулума или из пузырьков комплекса Гольджи. В меристематических клетках растений вначале возникает много мелких вакуолей. Увеличиваясь, они сливаются в центральную вакуоль, которая занимает до 70—90% объема клетки и может быть пронизана тяжами цитоплазмы.
Вакуоль в растительной клетке: 1 — вакуоль; 2 — цитопяаз-матические тяжи; 3 — ядро; 4 — хлоропласты. Содержимое вакуолей —клеточный сок. Он представляет собой водный раствор различных неорганических и органических веществ. Большинство из них являются продуктами метаболизма протопласта, которые могут появляться и исчезать в различные периоды жизни клетки. Химический состав и концентрация клеточного сока очень изменчивы и зависят от вида растений, органа, ткани и состояния клетки. В клеточном соке содержатся соли, сахара (прежде всего сахароза, глюкоза, фруктоза), органические кислоты (яблочная, лимонная, щавелевая, уксусная и др.), аминокислоты, белки. Эти вещества являются промежуточными продуктами метаболизма, временно выведенными из обмена веществ клетки в вакуоль. Они являются запасными веществами клетки. Помимо запасных веществ, которые могут вторично использоваться в метаболизме, клеточный сок содержит фенолы, танины (дубильные вещества), алкалоиды, антоцианы, которые выводятся из обмена в вакуоль и таким путем изолируются от цитоплазмы. Танины особенно часто встречаются в клеточном соке (а также в цитоплазме и оболочках) клеток листьев, коры, древесины, незрелых плодов и семенных оболочек. Алкалоиды присутствуют, например, в семенах кофе (кофеин), плодах мака (морфин) и белены (атропин), стеблях и листьях люпина (люпинин) и др. Считается, что танины с их вяжущим вкусом, алкалоиды и токсичные полифенолы выполняют защитную функцию: их ядовитый (чаще горький) вкус и неприятный запах отталкивают растительноядных животных, что предотвращает поедание этих растений. В вакуолях также часто накапливаются конечные продукты жизнедеятельности клеток (отходы). Таким веществом для клеток растений является щавелевокислый кальций, который откладывается в вакуолях в виде кристаллов различной формы. В клеточном соке многих растений содержатся пигменты, придающие клеточному соку разнообразную окраску. Пигменты и определяют окраску венчиков цветков, плодов, почек и листьев, а также корнеплодов некоторых растений (например, свеклы). Клеточный сок некоторых растений содержит физиологически активные вещества — фитогормоны (регуляторы роста), фитонциды, ферменты. В последнем случае вакуоли действуют как лизосомы. После гибели клетки мембрана вакуоли теряет избирательную проницаемость, и ферменты, высвобождаясь из нее, вызывают автолиз клетки. Функции вакуолей следующие: 1. Вакуоли играют главную роль в поглощении воды растительными клетками. Вода путем осмоса через ее мембрану поступает в вакуоль, клеточный сок которой является более концентрированным, чем цитоплазма, и оказывает давление на цитоплазму, а следовательно, и на оболочку клетки. В результате в клетке развивается тургорное давление, определяющее относительную жесткость растительных клеток и обусловливающее растяжение клеток во время их роста. 2. В запасающих тканях растений вместо одной центральной часто бывает несколько вакуолей, в которых скапливаются запасные питательные вещества (жиры, белки). Сократительные (пульсирующие) вакуоли служат для осмотической регуляции, прежде всего, у пресноводных простейших, так как в их клетки путем осмоса непрерывно поступает вода из окружающего гипотонического раствора (концентрация веществ в речной или озерной воде значительно ниже, чем концентрация веществ в клетках простейших). Сократительные вакуоли поглощают избыток воды и затем выводят ее наружу путем сокращений.
Содержимое вакуоли — клеточный сок — это водный раствор очень многих веществ: сахаров, аминокислот, других органических кислот, пигментов (красящих веществ), витаминов, дубильных веществ, алкалоидов, гликозидов, неорганических солей (нитратов, фосфатов, хлоридов), иногда — белков.
Кристаллы щавелевокислого кальция в вакуолях клеток. Все эти вещества — продукты жизнедеятельности клетки. Одни из них хранятся в вакуолярном (клеточном) соке в качестве запасных веществ и со временем вновь поступают в цитоплазму для использования. Другие являются отбросами обмена веществ, выведенными прочь из цитоплазмы. Так, в вакуоль выводится щавелевая кислота; в вакуолярном соке часто откладываются кристаллы щавелевокислого кальция — иногда в форме одиночных кристаллов, в других случаях в виде конгломерата кристаллов этой соли — многогранных (друзы) или игольчатых (рафиды). Клеточный центр(центросома) митотический центр, постоянная структура почти всех животных и некоторых растительных клеток, определяет полюса делящейся клетки. Клеточный центр обычно состоит из двух центриолей — плотных гранул размером 0,2—0,8 мкм, расположенных под прямым углом друг к другу. При образовании митотического аппарата центриоли расходятся к полюсам клетки, определяя ориентировку веретена деления клетки. В ходе развития организма изменяются как положение Клеточный центр в клетках, так и форма его. При делении клетки каждая из дочерних клеток получает пару центриолей. Процесс их удвоения происходит чаще в конце предыдущего клеточного деления. Возникновение ряда патологических форм деления клетки связано с ненормальным делением клеточный центр Центриоли представляют собой полые цилиндры длиной не более 0,5 мкм. Они располагаются парами перпендикулярно одна к другой. Каждая центриоль построена из девяти триплетов микротрубочек. Центриолям по структуре идентичны базальные тельца, которые всегда обнаруживаются в основании жгутиков и ресничек. Базальные тельца образуются путем удвоения центриолей. Базальные тельца, как и центриоли, являются центрами организации микротрубочек, входящих в состав жгутиков и ресничек.
Ядро У подавляющего большинства растений (исключение составляют прокариотические организмы) в каждой живой клетке имеется ядро или несколько ядер. Ядро всегда лежит в цитоплазме. Форма ядра может быть различной — округлой, овальной, сильно вытянутой, неправильно-многолопастной. В некоторых клетках контуры ядра меняются в ходе его функционирования, причем на его поверхности образуются лопасти различной величины. Размеры ядер неодинаковы и в клетках разных растений, и в разных клетках одного и того же растения. Относительно крупные ядра бывают в молодых, меристематических клетках, в которых они могут занимать до 3/4 объема всей клетки. Относительные, а иногда и абсолютные размеры ядер в развитых клетках значительно меньше, чем в молодых.
.От цитоплазмы ядро отделено ядерной оболочкой, образованной за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жёсткой белковой структурой, образованной белками-ламинами , к которой прикреплены нити хромосомной ДНК. Ламины прикрепляются к внутренней мембране ядерной оболочки при помощи заякоренных в ней трансмембранных белков — рецепторов ламинов.В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой. Пора не является дыркой в ядре, а имеет сложную структуру, организованную несколькими десятками специализированных белков — нуклеопоринов. Внешняя из двух мембран оболочки дает выросты, непосредственно переходящие в стенки эндоплазматической сети цитоплазмы. И поры и прямая связь эндоплазматической сети с околоядерным пространством обеспечивают тесный контакт между ядром и цитоплазмой Содержимое ядра — зернистое основное вещество (ядерный сок, или нуклеоплазма), в котором помещаются более плотные структуры — хромосомы и ядрышко. Ядрышко представляет собой аппарат синтеза материала рибосом и место их сборки из этого материала. Ядрышко находится внутри ядра, и не имеет собственной мембранной оболочки, однако хорошо различимо под световым и электронным микроскопом. Основной функцией ядрышка является синтез рибосом. В геноме клетки имеются специальные участки, так называемые ядрышковые организаторы, содержащие гены рибосомной РНК (рРНК), вокруг которых и формируются ядрышки. В ядрышке происходит синтез рРНК РНК полимеразой I, ее созревание, сборка рибосомных субчастиц. В ядрышке локализуются белки, принимающие участие в этих процессах. Некоторые из этих белков имеют специальную последовательность — сигнал ядрышковой локализации. Следует отметить, самая высокая концентрация белка в клетке наблюдается именно в ядрышке. В этих структурах было локализовано около 600 видов различных белков, причем считается, что лишь небольшая их часть действительно необходима для осуществления ядрышковых функций, а остальные попадают туда неспецифически. Под электронным микроскопом в ядрышке выделяют несколько субкомпартментов. Так называемые Фибриллярные центры окружены участками плотного фибриллярного компонента, где и происходит синтез рРНК. Снаружи от плотного фибриллярного компонента расположен гранулярный компонент, представляющий собой скопление созревающих рибосомных субчастиц. Наследственная информация представлена хроматином. Хроматином называют молекулы хромосомной ДНК в комплексе со специфическими белками. Основную массу составляют «белки хранения», так называемые гистоны. Из этих белков построены нуклеосомы, структуры на которые намотаны нити молекул ДНК. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из белков четырех типов: H2A, H2B, H3 и H4. В одну нуклеосому входят по два белка каждого типа — всего восемь белков. Гистон H1, более крупный чем другие гистоны, связывается с ДНК в месте ее входа на нуклеосому. Нуклеосома вместе с H1 называется хроматосомой. Нить ДНК с нуклеосомами образует нерегулярную соленоид-подобную структуру толщиной около 30 нанометров, так называемую 30 нм фибриллу. Дальнейшая упаковка этой фибриллы может иметь различную плотность. Если хроматин упакован плотно, его называют конденсированным или гетерохроматином, он хорошо видим под микроскопом. ДНК, находящаяся в гетерохроматине, не транскрибируется, обычно это состояние характерно для незначащих или молчащих участков. В интерфазе гетерохроматин обычно располагается по периферии ядра (пристеночный гетерохроматин). Полная конденсация хромосом происходит перед делением клетки. Если хроматин упакован неплотно, его называют эу- или интерхроматином(эухроматин) Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется наличием транскрипционной активности. Плотность упаковки хроматина во многом определяется модификациями гистонов — ацетилированием и фосфориллированием. Считается, что в ядре существуют так называемые функциональные домены хроматина(ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную территорию
Внутренняя структура (А) и внешний вид (5) хромосомы. 1 - хроматиды (половины хромосомы, на которые она разделяется продольно во время деления клетки), 2 - хромонемы (слабо спирализованные парные нити ДНК, слагающие основу хромосомы), 3 - белковый матрикс хромосомы, 4 - первичная перетяжка с центромерой, 5 - спутник хромосомы.
Организация хроматина в ядре Хроматин – это основное вещество хромосом эукариот, который представляет собой комплекс ДНК с белками (гистоновыми и негистоновыми). Результатом взаимодействия ДНК с белками в составе хроматина является ее компактизация. Суммарная длина ДНК, заключенной в ядре клеток человека, приближается к 1 м, тогда как средний диаметр ядра составляет 10 мкм. Длина молекулы ДНК, заключенной в одной хромосоме человека, в среднем равняется ~4 см. В то же время длина метафазной хромосомы составляет 4 мкм. Следовательно, ДНК метафазных хромосом человека компактизована по длине, по крайней мере, в 104 раз. Степень компактизации ДНК в интерфазных ядрах значительно ниже и неравномерна в отдельных генетических локусах. Эухроматин характеризуется меньшей компактизацией ДНК, и в нем главным образом локализуются активно экспрессирующиеся гены. Гетерохроматин представляет собой наиболее компактные участки хроматина, где локализуются неактивные гены. Гетерохроматизация определенных участков хромосом часто сопровождается подавлением транскрипции имеющихся в них генов. В процесс гетерохроматизации могут быть вовлечены протяженные участки хромосом и даже целые хромосомы (например, Х-хромосома у женщин).
|
|||||||||
|
|
Последнее изменение этой страницы: 2017-02-19; просмотров: 478; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.141.202.187 (0.08 с.) |