Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Биосинтез нуклеиновых кислот и белков. Основы молекулярной генетики
20. Механизм матричных биосинтезов: репликация ДНК. Повреждения ДНК, повреждающие факторы. Репарация ДНК. Последствия повреждений ДНК. 21. Механизмы матричных биосинтезов: транскрипция, обратная транскрипция. Ингибиторы процессов. 22. Биосинтез белков. Биологический код. Последовательность этапов в синтезе полипептидной цепи. Ингибиторы трансляции (интерфероны и др.). Посттрансляционная модификация белка. 23. Клеточная дифференцировка и онтогенез как результат регуляции активности генов. Доказательство идентичности первичной структуры ДНК в разных типах клеток одного организма. 24. Молекулярные механизмы генетической изменчивости: молекулярные мутации. Влияние условий среды. Наследственные болезни (биохимические основы и механизмы развития патологии). Методы диагностики и коррекции. 25. Пути синтеза пуриновых и пиримидиновых нуклеотидов, ферменты, регуляция процесса. Оротацидурия 26. Катаболизм экзогенных и эндогенных нуклеиновых кислот в желудочно-кишечном тракте. Нуклеазы слюны, их защитное действие. 27. Катаболизм пуриновых нуклеотидов. Образование и выделение мочевой кислоты. Гиперурикемия, причины и следствия. 28. Патология обмена нуклеотидов, возможные причины, формирование патологии (подагра, синдром Леша-Нихана).
20) Процесс удвоения хромосом называют репликацией.Репликацию можно разделить на 4 этапа: образование репликативной вилки (инициация), синтез новых цепей (элонгация), исключение праймеров, завершение синтеза двух дочерних цепей ДНК (терминация).Синтез ДНК у эукариотов происходит в S-фазу клеточного цикла. Инициацию репликации регулируют специфические сигнальные белковые молекулы - факторы роста. Факторы роста связываются рецепторами мембран клеток, которые передают сигнал, побуждающий клетку к началу репликации.Синтез новых одноцепочечных молекул ДНК может произойти только при расхождении родительских цепей. В определённом сайте (точка начала репликации) происходит локальная денатурация ДНК, цепи расходятся и образуются две репликативные вилки, движущиеся в противоположных направлениях. Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами (рис. 4-16). Субстратами и источниками энергии для синтеза продукта служат 4 макроэргических соединения - дезоксирибонуклеозидтрифосфаты дАТФ, дГТФ, дЦТФ и дТТФ, для активации которых необходимы ионы магния. Нейтрализуя отрицательный заряд нуклеотидов, они повышают их реакционную способность. Ферменты проявляют каталитическую активность только в присутствии предварительно раскрученной матричной двухцепочечной ДНК. Синтез цепей ДНК происходит в направлении 5'→3' растущей цепи, т.е. очередной нуклеотид присоединяется к свободному 3'-ОН-концу предшествующего нуклеотид-ного остатка. Синтезируемая цепь всегда антипараллельна матричной цепи. В ходе репликации образуются 2 дочерние цепи, представляющие собой копии матричных цепей.
Индуцируемые повреждения возникают в ДНК в результате воздействия разнообразных мутагенных факторов как радиационной, так и химической природы. Процесс, позволяющий живым организмам восстанавливать повреждения, возникающие в ДНК, называют репарацией. Все репарационные механизмы основаны на том, что ДНК - двухцепочечная молекула, т.е. в клетке есть 2 копии генетической информации. Если нуклеотидная последовательность одной из двух цепей оказывается повреждённой (изменённой), информацию можно восстановить, так как вторая (комплементарная) цепь сохранена. Процесс репарации происходит в несколько этапов. На первом этапе выявляется нарушение комплементарности цепей ДНК. В ходе второго этапа некомплементарный нуклеотид или только основание устраняется, на третьем и четвёртом этапах идёт восстановление целостности цепи по принципу комплементарности. Однако в зависимости от типа повреждения количество этапов и ферментов, участвующих в его устранении, может быть разным. Очень редко происходят повреждения, затрагивающие обе цепи ДНК, т.е. нарушения структуры нуклеотидов комплементарной пары. Такие повреждения в половых клетках не репарируются, так как для осуществления сложной репарации с участием гомологичной рекомбинации требуется наличие диплоидного набора хромосом. 21) Транскрипция - первая стадия реализации генетической информации в клетке. В ходе процесса образуются молекулы мРНК, служащие матрицей для синтеза белков, а также транспортные, рибосомальные и другие виды молекул РНК, выполняющие структурные, адапторные и каталитические функции
В процессе транскрипции различают 3 стадии: инициацию, элонгацию и терминацию. Инициация.Активация промотора происходит с помощью большого белка - ТАТА-фактора, называемого так потому, что он взаимодействует со специфической последовательностью нуклеотидов промотора - ТАТААА- (ТАТА-бокс) (рис. 4-29). Присоединение ТАТА-фактора облегчает взаимодействие промотора с РНК-полимеразой. Факторы инициации вызывают изменение кон-формации РНК-полимеразы и обеспечивают раскручивание примерно одного витка спирали ДНК, т.е. образуется транскрипционная вилка,в которой матрица доступна для инициации синтеза цепи РНК (рис. 4-30). После того как синтезирован олигонуклеотид из 8-10 нуклеотидных остатков, σ-субъединица отделяется от РНК-полимеразы, а вместо неё к молекуле фермента присоединяются несколько факторов элонгации. Элонгация Факторы элонгации повышают активность РНК-полимеразы и облегчают расхождение цепей ДНК. Синтез молекулы РНК идёт от 5'- к З'-концу комплементарно матричной цепи ДНК. На стадии элонгации, в области транскрипционной вилки, одновременно разделены примерно 18 нуклеотидных пар ДНК. Растущий конец цепи РНК образует временную гибридную спираль, около 12 пар нуклеотидных остатков, с матричной цепью ДНК. По мере продвижения РНК-полимеразы по матрице в направлении от 3'- к 5'-концу впереди неё происходит расхождение, а позади - восстановление двойной спирали ДНК. Терминация Раскручивание двойной спирали ДНК в области сайта терминации делает его доступным для фактора терминации. Завершается синтез РНК в строго определенных участках матрицы - терминаторах (сайты терминации). Фактор терминации облегчает отделение первичного транскрипта (пре-мРНК), комплементарного матрице, и РНК-полимеразы от матрицы. РНК-полимераза может вступить в следующий цикл транскрипции после присоединения субъединицы σ. К ингибиторам матричных синтезов, оказывающим противобактериальное действие, относят вещества, блокирующие синтез РНК или белка. В эту группу входит широко применяемый в клинике рифампицин, получаемый на основе природного антибиотика рифамицина. Антибиотики из семейства рифамицинов ингибируют только бактериальную ДНК-зависимую РНК-полимеразу, связываясь с β-субъединицей фермента и препятствуя инициации транскрипции (рис. 4-44). Их применяют для лечения туберкулёза, так как эти препараты не влияют на работу ядерных РНК-полимераз эукариотических клеток. Однако они могут ингибировать синтез митохондриальных РНК, хотя дозы препарата, при которых блокируется образование митохондриальных РНК, выше тех, что используют в лечении инфекционного заболевания. Ингибиторы транскрипции Рифамицины Связываются с бактериальной РНК-полимеразой и препятствуют началу транскрипции обратная транскрипция - это синтез ДНК по матрице РНК.Обратная транскрипция — это процесс образования двуцепочечной ДНК на матрице одноцепочечной РНК. Данный процесс называется обратной транскрипцией, так как передача генетической информации при этом происходит в «обратном», относительно транскрипции, направлении
Ингибиторы обратной транскриптазы делятся на нуклеозидные (НИОТ), нуклеотидные (НтИОТ) и ненуклеозидные (ННИОТ). НИОТ мешают вирусу тем, что поставляют негодный строительный материал для создания вирусной ДНК - неправильные нуклеозиды (рис. 30А). В результате процесса обратной транскрипции при наличие аномального строительного материала образуется испорченный продукт. На сходном принципе основано действие нуклеотидного ингибитора обратной транскриптазы, только он поставляет "бракованные" нуклеотиды, а не нуклеозиды. Так как ферменты клетки для синтеза своей ДНК используют те же стройматериалы, что и вирусная обратная транскриптаза, то главная задача при подборе и синтезе аналогов нуклеозидов и нуклеотидов с антиретровирусной активностью - максимальное их сродство с вирусной обратной транскриптазой и минимальное - с клеточными ДНК-полимеразами. 22) Биосинтез белка- сложный многостадийный процесс синтеза полипиптидной цепи из аминокислот, происходящий на рибосомах с участием молекул мРНК и тРНК. Этот процесс требует значительных затрат энергий. Биологический код(генет. код), свойственный всем живым ор-м сопсоб кодиров-я аминокислот последов-ти белков при помощи послед-ти нукл. Трансляция-синтез полипеп цепей. 1) инициация-происходит образ-е целбной рибосомы, присоед мРНК и устан-е первой аминок-ты.2) элонгация-ко 2 кодону присоед еще одна аминоацил-тРНК. М/д карбокс группой группой первой аминкты и амино группой второй образ пепт связь. После это 1 аминкта отсоедин-ся от своей тРНК.3) терминация-синтез полипеп цепей идет до тех пор пока рибосома не достигнет одного из 3 стоп кодонов. В этот момент белков цепь отделится, а рибосома диссоциирует на суб единицы. Посттрансляц модификац-это ковалент химич модификация белка после его синтеза на рибосоме. Для некот белков посттрансляц модификация оказыв-ся заверш этапом биосинтза. 23.) Клеточн дифференцировка- пр-сс возникн-я разл м/д первоначал и однород клетками, в ходе кот. обр-ся специализир клетки, ткани и ор-ны, способн вып-ть в ор-ме опр-е функц. таким образом,дифференцировка лежит в основе индивид разв-я многоклет ор-ов от оплодотвор до взросл особи. 24) Генет изм-ть- изм-ть обусловлена взаимодействием и различным проявлением генет факторов. В основе генной инженерии может лежать мутация, рекомбинация, изм-ть, привод к разл сочетаниям аллелей в пр-ссе случ расхожд-я хромосом при мейозе. *Мутация- изм-е в генотипе, кот стабилно наслед-ся. Бывают мутац: спонтанные, индуцированные. * Диаг-ка наслед болезней. Цитогенет -выяв-е хромосом болезней. Биохим-некот бол-ни хар-ся биохим изм-ми кот св-ны с наруш опред метабол пути. Молекулярно-генет-й-самый совр метод выявл генет изм-й.
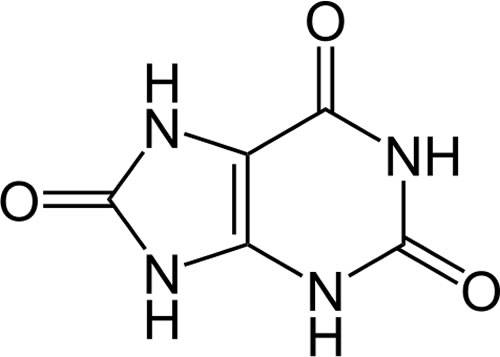
25) Синтез пуриновых нуклеотидов- слож многостад пр-сс. на первой стадии происх построение пуринового кольца инозинмонофосфата. На второй ст-ии ИМФ преобр-ся в оцениловые и гуальнов рибо и дезоксирибонуклеотиды. Стр-ра пиримид кольа проще и путь биосинтеза пиримидинов короче чем у пуринов. Амидн N глутамина и диоксид С обесп атомы 2 и 3 кольца пиримидина после преобраз-я в карбамоилфосфат. Др 4 атома кольца происх из аспартата. также как и в случае с пуринов нуклеотидами, углерод часть постав-ся ФРПФ(фосфорибозилпирофосфат) оротоцидурия- наслед болезнь обусловл недост-ть ферментов, перевод ортовую к-ту в цитидилову. Хар-ся тяжелой анемией и отлож-ем в тканях и ор-ах кристаллов ортовой к-т; наслед-ся аутосомно по рецесив признаку. 26) Нуклеаза слюны участв в деград нуклеин к-т, вирусов и поэтому играет ведущ роль в защите ор-ма от вирус инфек-ии. Нуклеин к-ты- природ высокомолек орг-я соед-я обеспечив хранение и передаччу наслед инфор Сущ 3 типа нукл к-т кот раз по стр-ю и ф-ции. одна сод рибозу друг содер дизоксирибозу. 27) Продуктом кат-ма пурин нуклотид явл-ся мочевая к-та. ее обр-е идет путем гидролит отщепления фосфат ост-ка от нукл-в. С помощью нуклеотидаз или фосфатаз, фосфрилаза N-гликозид связи нуклеозидов и оксиление азот основ-й. Мочев к-та обр-ся в рез-те распада пуриновых нуклеотидов . Гиперуренемия- повыш сод моч к-ты в крови. Бол-нь выз-я ускор обр-ем мочев к-ты из за уч-я пурина в обмене в.в. или из за ослабл работы почек, или из за повыш содер фруктозы в пище. 28) т.к. нуклеотиды не имею специализ конеч про-ов обмена поэтому при сост хар-ся избыт синтезом как правило нет выраж клинич признаков. При тормож синтеза обуславл-ся недостатком в ор-ме фолиевой к-ты идет одновр и нар-е синтеза пуринов нуклеотидов, что прояв-ся в виде нар-я синтеза нуклеин к-т с разв-ем той или иной форм анемии. Подагра-гетерог по происх заб-е кот хар-ся отлож-ем кристаллов мочев к-ты в разл тк-ях ор-ма. Синдром Лёша-Нихана-наслед заб-е хар-ся ув-ем синтеза мочев к-ты, след-но обр-ся много мочев к-ты.
|
||||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 294; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.133.108.241 (0.011 с.) |