
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Производные зародышевых листков. Органогенез
20. Развитие производных эктодермы (развитие нервной системы и кожных покровов) 21. Развитие производных эктодермы (развитие органов зрения, слуха, вкуса и обоняния) 22. Развитие производных энтодермы 23. Развитие производных мезодермы (сомиты, нефрогонотомы, хорда) 24. Развитие производных мезодермы (листки спланхнотомов и мезенхима) 25. Нервный гребень как «четвёртый зародышевый листок». Производные нервного гребня 26. Явление эмбриональной индукции в механизмах онтогенеза: опыты Г. Шпемана 27. Природа индуцирующего воздействия. Явления первичной, вторичной и третичной индукции 28. Генетические механизмы детерминации и дифференцировки 29. Развитие ланцетника (тип яйцеклетки, дробление, тип бластулы, гаструляция, нейруляция, гистогенез, органогенез) 30. Развитие амфибий (тип яйцеклетки, дробление, тип бластулы, гаструляция, нейруляция, гистогенез, органогенез) 31. Развитие птиц (тип яйцеклетки, дробление, тип бластулы, гаструляция, нейруляция, гистогенез, органогенез) 32. Особенности эмбриогенеза млекопитающих 33. Эмбриональное развитие человека 34. Образование и функции плаценты. Значение в эмбриогенезе плацентарных млекопитающих 35. Гистологическая классификация плацент.
Биология индивидуального развития 1. Общая характеристика гаметогенеза У высших млекопитающих существует половое размножение - происходит с помощью половых клеток. Бесполое (вегетативное) размножение –размножение без участия половых клеток, новый организм развивается из соматических клеток, сохраняющих свойства модифицированных клеток. Основные отличительные признаки половых клеток от соматических: 1. Половым клеткам на определенном этапе развития свойственен специфический, только им присущий, вид деления – мейоз. 2. Имеют половинный набор хромосом от характерного для соматических клеток данного вида. 3. У половых клеток изменен ядерно-цитоплазматический индекс (соотношение ядра и цитоплазмы). 4. Интенсивный рост женских половых клеток обеспечивается последовательным развитием запаса цитоплазматических компонентов, в т. ч. нуклеиновых кислот и специфического для яйцеклеток включения – желтка. 5. Яйцеклетки окружены более сложными оболочками, чем соматические клетки.
6. У зрелых половых клеток процессы диссимиляции и ассимиляции минимальны, т. е. заторможен обмен веществ. 7. Для большинства сперматозоидов свойственно активное перемещение во внешней среде. 8. Половые клетки – это высоко дифференцированные клетки, выполняющие специфические функции. Гаметогенез Появление половых клеток и гаметогенез являются следствием роста, возникает эпигенетически. Их возникновение соответствует особому физиологическому состоянию, которое в норме приобретается развивающимся организмом. Прогенез – развитие половых клеток – происходит в половых железах. Наступившее гаметогенное физиологическое состояние организма распространяется на все ткани, в особенности на те из них, в которых находятся гоноциты. Сами по себе гоноциты сексуально индифферентны, внутри них не содержится факторов, детерминирующих пол гамет.Детерминация пола гамет исходит из тканей, в которых гаметогенные клетки находятся и дифференцируются. Эта лабильность половых клеток снижается у более специализированных форм, где пол детерминирован более жестко. Различают нескольких форм гаметогенеза: 1. Диффузный гаметогенез (у губок) 2. Локализованный гаметогенез - развитие половых клеток происходит в гонадах: в семенниках или яичниках (у большинства животных) 3. Солитарный гаметогенез - развитие и питание половых клеток осуществляется без участия специализированных вспомогательных элементов (кишечнополостные, черви) 4. Алиментарный гаметогенез - в развитии половых клеток участвуют специализированные клетки, например, трофоциты насекомых, фолликулярный эпителий гонад позвоночных. · Диффузный и локализованный гаметогенез может происходить как по солитарному, так и по алиментарному типу. Суть гаметогенеза состоит в образовании из диплоидных стволовых предшественников половых клеток, высокодифференцированных клеток – сперматозоидов и яйцеклеток – с гаплоидным набором хромосом.
2. Сперматогенез В извитых канальцах происходит сперматогенез - образование сперматозоидов из стволовых сперматогенных клеток. Эти канальцы содержатся в дольках по 1-4 канальца на дольку; всего в яичке 300-450 канальцев. В стенке канальца - два элемента: собственная оболочка - находится с наружной стороны стенки и сперматогенный "эпителий" - обращён к просвету канальца. Между канальцами находится рыхлая соединительная ткань - интерстиций, в её составе: сосуды, обычные соединительнотканные элементы и интерстициальные клетки (клетки Лейдига, или гландулоциты), продуцирующие мужские половые гормоны (тестостерон и его производные). В собственную оболочку семенного канальца входят (изнутри кнаружи) следующие компоненты:
1. Базальный слой: базальная мембрана миоидных клеток, 2. Миоидный слой: состоит из клеток, похожих на гладкие миоциты, но всё же отличных от них; клетки плотно прилегают друг к другу, образуя соединения, похожие на плотные; 3. Волокнистый слой: включает 2 слоя: наружный неклеточный слой и слой фибробластоподобных клеток (самый наружный). Таким образом, в собственной оболочке семенного канальца чередуются (изнутри кнаружи) два неклеточных и два клеточных слоя. Данная оболочка является одним из компонентов гематотестикулярного барьера: через неё не проходят иммунокомпетентные клетки. Сперматогенный "эпителий" лежит на базальной мембране и тоже включает два типа клеток: собственно эпителиальные (играющие вспомогательную роль) и сперматогенные клетки на разных стадиях созревания. Развитие сперматозоидов начинается в период пренатального развития при закладке генеративных тканей, затем возобновляется в период наступления половозрелости и продолжается до старости. В результате сперматогенеза возникают клетки – сперматозоиды, содержащие X- и Y-хромосому. Сперматогенные клетки: 1. Сперматогонии – стволовые (делятся очень редко, прилежат к базальной мембране) и вступившие в сперматогенез (активно делятся путём митоза, лежат вблизи от базальной мембраны); 2. Сперматоциты находятся в состоянии мейоза, клетки - крупные, в ядрах хорошо выражен рисунок хроматина, несколько удалены от базальной мембраны; 3. Сперматиды - ранние (непосредственные продукты мейоза: клетки гаплоидные, небольшого размера, округлой формы (как и все предыдущие клетки), со светлым ядром; находятся в средних слоях "эпителия") и поздние (образуются в результате созревания ранних сперматид: по размеру - мелкие, с плотными ядрами, а также со жгутиками (хвостами); находятся в слое, прилегающем к просвету канальцев); 4. Сперматозоиды близки по морфологии к поздним сперматидам, но лежат в просвете канальцев. Из этого описания следует, что по мере созревания сперматогенные клетки постепенно перемещаются вдоль клеток Сертоли - от базальной мембраны к просвету канальца. Они соответствуют четырём этапам сперматогенеза: 1 Период размножения 2 Период роста 3 Период созревания – мейоз 4 Период формирования 1.
2. Рост. Фаза роста выделяется условно в мужском организме, т. к. не связана с накоплением питательных веществ.Её часто объединяют с третьей фазой созревания в одну – мейотическую – фазу. На этой стадии их называют сперматоцитами I порядка (или цитами I). В мейотической фазе сперматоцит I порядка проходит длинную профазу I деления мейоза, которая у человека продолжается около двух суток. Рост характеризуется увеличением объема сперматоцита в 4 раза. 3. Созревание (мейоз). В сперматогенезе профаза непосредственно продолжается в последующие стадии I деления мейоза. В мейозе сперматоциты I проходят два процесса клеточного деления. В первом делении (редукционном) происходит уменьшение числа хромосом (редукция). В результате из одного цита I возникает две равновеликие клетки – сперматоциты IIпорядка, или циты II. Затем наступает второе деление созревания. Оно протекает как обычный соматический митоз, но при гаплоидном числе хромосом. Такое деление называется эквационным («эквацио» – равенство), так как образуются две тождественные, т.е. полностью равноценные клетки, которые называются сперматидами. Сперматиды отличаются друг от друга набором хромосом: они все содержат по 22 аутосомы, но половина клеток содержит Х-хромосому, а другая половина – Y-хромосому. Аутосомы отличаются между собой и от родительских различным сочетанием аллелей, поскольку произошел обмен во время кроссинговера. 4. Формирование. Округлая сперматида приобретает форму зрелой мужской половой клетки: у нее вырастает жгутик, уплотняется ядро, образуется оболочка. В результате всего процесса сперматогенеза из каждой исходной недифференцированной сперматогонии получается 4 зрелых половых клетки, содержащих по гаплоидному набору хромосом. · Сперматозоиды состоят из 4 частей: головки, шейки, средней части и хвоста. · Головка различна по форме - от округлой до овально-веретеновидной. Основной компонент головки - клеточное ядро с гаплоидным набором конденсированных хромосом. Плотность упаковки хромосом велика, в таком конденсированном состоянии генетический материал более защищён от повреждений при прохождении сперматозоида через оболочки яйцеклетки.Спереди большая часть ядра, как двойной шапочкой, покрыта акросомой - уплощённый пузырёк, происходящий из комплекса Гольджи, содержит ферменты, необходимые для проникновения сперматозоида в яйцеклетку (коллагеназу, гиалуронидазу, акрозин, кислую фосфатазу и др.), стимулирует и активирует яйцо.
· Вся головка сперматозоида в целом, как и его хвост, окружена плазматической мембраной. В области головки эта мембрана содержит специальные белки - одни способствуют направленному движению сперматозоида к яйцеклетке, другие белки участвуют в связывании с яйцеклеткой. · В шейке содержатся проксимальная и дистальная центриоли. Проксимальная центриоль играет роль в делении оплодотворенного яйца. Дистальная центриоль участвует в образовании осевой нити хвоста. Осевая нить – главный элемент в хвосте сперматозоида, который обеспечивает его подвижность. · В средней части находится митохондрии, которые обеспечивают метаболическую и энергетическую активность сперматозоида. · В хвосте различают 4 части: шейку (I), промежуточную часть (II), основную часть (III), концевую часть (IV). Короткая шейка содержит 2 центриоли - полые цилиндры из микротрубочек. От одной из центриолей начинается аксонема хвоста - идёт вдоль оси до самого конца хвоста. Она имеет обычное для всех жгутиков и ресничек строение, т.е. образована микротрубочками. Микротрубочки взаимодействуют с помощью динеиновых ручек; за счёт этого происходит скольжение соседних дуплетов друг относительно друга, приводящее к биению хвоста. В промежуточной части вокруг аксонемы имеется митохондриальная "оболочка"- образована митохондриями, расположенными по спирали, витки которой плотно прилегают друг к другу. · В организме мужчины сперматозоиды находятся в неактивном состоянии, движения жгутиков у них незначительны. Перемещение сперматозоидов по половым путям мужчины происходит пассивно за счет перистальтических сокращений мышц протоков и биения ресничек клеток стенок протоков. Сперматозоиды приобретают активность после эякуляции за счет воздействия на них ферментов простатического сока. Движение сперматозоидов по половым путям женщины является самостоятельным и осуществляется против движения жидкости. Для осуществления оплодотворения сперматозоидам необходимо преодолеть путь длиной около 20 см (цервикальный канал — около 2 см, полость матки — около 5 см,фаллопиева труба — около 12 см). Направление движения сперматозоид определяет, воспринимая pH окружающей среды. Он движется по направлению уменьшения кислотности. Среда матки оказывает на сперматозоиды активирующее действие, их подвижность значительно возрастает. Это явление получило название «капацитация». Из матки сперматозоиды направляются в фаллопиевы трубы, направление к которым и внутри которых сперматозоиды определяют по току жидкости. · У человека сперматогенез продолжается до глубокой старости. Продолжительность полного сперматогенеза у мужчин составляет примерно 73—75 дней. 3. Гормональная регуляция сперматогенеза Синтез и секреция мужских половых гормонов регулируется гипоталамо-гипофизарной системой по механизму отрицательной обратной связи. Секреция лютеинизирубщего (ЛГ) и фолликулостимулирующего (ФСГ) стимулируется гонадотропин-рилизинг гормоном. В препубертатный период секреция андрогенов подавляет по механизму отрицательной обратной связи секрецию гонадотропина до начала пубертатного периода, когда гипофизарные клетки становятся менее чувствительными к ингибирующему действию циркулирующих в крови андрогенов. Эта потеря чувствительности приводит к циклически импульсному освобождению ЛГ и ФСГ. ЛГ, связываясь с рецепторами клеток Лейдига, стимулирует образование тестостерона интерстициальными клетками Лейдига, а ФСГ, связываясь с рецепторами клеток Сертоли в семенниках, стимулирует сперматогенез.
В этих клетках находятся рецепторы ФСГ, через которые активируется выработка андрогенсвязывающего белка (АСБ). Комплекс тестостерона с этим белком оказывает влияние на сперматогенез.Тестостерон замыкает отрицательную обратную связь на уровне гипофиза и гипоталамуса, уменьшая частоту секреторных импульсов ЛГ. Торможение секреции ФСГ аденогипофизом происходит под действием белка ингибина, вырабатываемого клетками Сертоли. ФСГ стимулирует синтез этого белка, который по механизму отрицательной обратной связи тормозит дальнейшую секрецию ФСГ. (Клетки Сертоли вырабатывают и белок активин, который обладает противоположным действием (механизм положительной обратной связи, на рисунке не показано)).
4. Оогенез Процесс развития женских половых клеток называется оогенезом. Количество образующихся яйцеклеток у млекопитающих на много порядков меньше, чем сперматозоидов.В результате оогенеза возникают клетки – ооциты (яйцеклетки), содержащие Х-хромосому. Оогенез включает 3 стадии: 1. Размножение. Попав в яичник, гоноциты становятся оогониями. Оогонии осуществляют период размножения, в ходе клеточного цикла синтезируют ДНК и митотически делятся. В женском организме фаза размножения инициируется и завершается еще в эмбриогенезе. К окончанию данной фазы в каждом яичнике плода заключено 6 – 7 млн. овогоний. 2. Рост. Период роста. Длится очень долго – недели, месяцы, годы. Различают 2 этапа – малый (медленный) рост – накопление веществ в цитоплазме и большой (быстрый) рост – вителлогенез – накопление питательного желтка. Малый рост происходит в эмбриогенезе, а большой рост – после полового созревания, в репродуктивном периоде. Глубокие изменения претерпевает ядро, оно набухает. Размеры клеток колоссально возрастают. Овоцит первого порядка вступает в период созревания, или мейоз. 3. Созревание. В овогенезе половые клетки останавливаются в стадии диакинеза под влиянием мейозингибирующей субстанции, и могут прибывать в ней несколько лет – стационарная стадия профазы I деления мейоза. Женские половые клетки выходят из стационарного состояния и продолжают своё развитие в разные периоды репродуктивного возраста. Многие из них погибают. Фактором, стимулирующим продолжение мейоза является мейозстимулирующая субстанция, вырабатываемая фолликулами яичников. Собственно мейоз включает в себя два последовательных деления созревания: 1. Редукционное. 2. Эквационное. Профаза I деления мейоза в сперматогенезе и оогенезе включает 5 стадий: 1. Лептотена. Реплицированные хромосомы деспирализуются и принимают вид длинных и спутанных нитей. 2. Зиготена. Гомологичные по отцовской и материнской линиям хромосом сближаются, соединяются попарно – конъюгируют, образуются биваленты. При конъюгации контактируют друг с другом строго гомологичные хромосомы. 3. Пахитена. Хромосомы обмениваются геномами и группами генов – кроссинговер. 4. Диплотена. Гомологичные хромосомы, образующие бивалент, отходят друг от друга, но между ними сохраняется связь в области центромеры благодаря хиазмам. 5. Диакинез. Заключительная стадия профазы I деления мейоза, характеризуется спирализацией и укорочением хромосом. Метафаза I деления мейоза. Тетрады хроматид выстраиваются по экватору. Распределение отцовских и материнских хромосом абсолютно случайно. Исчезает ядерная мембрана. Возникает веретено деления. В процессе анафазы и телофазы в каждую дочернюю клетку расходится по одной гомологичной двойной хромосоме от каждого бивалента. Это происходит потому, что при I делении мейоза центромера каждого бивалента не расцепляется, и каждый бивалент имеет собственные кинетохоры, которые контактируют с веретеном деления. Таким образом, каждая дочерняя клетка получает гаплоидный набор хромосом, в связи с чем деление названо редукционным. При этом делении одно ядро увлекает с собой большую часть цитоплазмы, а на долю другого остается лишь незначительная ее часть. Поэтому образуется только одна полноценная клетка – овоцит второго порядка, и вторая крошечная – направительное, или редукционное тельце, которое может делиться на два. II деление мейоза Поскольку первому делению созревания предшествовала S-фаза, каждая из разошедшихся хромосом содержит двойное количество ДНК (две хроматиды). Эти генетически идентичные хроматиды и расходятся по сестринским клеткам во втором делении созревания, которое является эквационным. При втором делении несимметричное распределение цитоплазмы повторяется и опять образуется одна крупная клетка – овотида и третье полярное тельце. Период формирования отсутствует. Кроме этого в периоде созревания существует 2 блока мейоза: от момента рождения до полового созревания – блок профазы первого деления мейоза. Этот блок снимается благодаря регулирующему действию гормонов ГГП оси, и второй блок – метафазы второго деления снимается в момент оплодотворения. Строение яйцеклеток 1. Для яйцеклеток характерна полярность. 2. Они имеют сложные яйцевые оболочки, которых нет у сперматозоидов. 3. Они имеют кортикальную оболочку, которая образуется при оплодотворении путем слияния кортикальных гранул с плазмолеммой. 4. У яйцеклеток отсутствует локомоторный аппарат. Редукционные тельца: при мейозе исходная тетраплоидная клетка (ооцит) подразделяется на четыре гаплоидные клетки. Но при этом образуется лишь одна яйцеклетка, приобретающая практически всю цитоплазму ооцита вместе с её запасными питательными веществами. Остальные же клетки - продукты мейоза - это т.н. редукционные тельца: они получают лишь необходимую часть хромосом и практически лишены цитоплазмы. Редукционные тельца часто остаются под оболочками, окружающими яйцеклетку. Условность термина "яйцеклетка": в покоящихся и созревающих фолликулах яичников содержатся не яйцеклетки, а их предшественники - ооциты I и затем Яичник снаружи покрыт однослойным плоским эпителием – мезотелием. Под ним находится белочная оболочка (плотная волокнистая СТ). Паренхима яичника подразделяется на корковое вещество и мозговое – узкую область в глубине яичника с сосудами и нервами. Главные компоненты - фолликулы - сосредоточены в корковом веществе. Каждый фолликул содержит ооцит на определенной стадии мейоза и фолликулярные клетки, имеющие эпителиальную природу, лежащие на базальной мембране. Фолликулогенез (созревание фолликулов) — непрерывный процесс, который, начавшись в антенатальном периоде, заканчивается лишь после наступления менопаузы. Большинство фолликулов гибнет на различной стадии развития в результате апоптоза («атрезия фолликулов»), только небольшая их часть проходит все стадии созревания и участвует в овуляции. К наступлению менопаузы происходит возрастное истощение запаса фолликулов в яичниках. В соответствии со стадией развития различают примордиальные, преантральные (первичные), антральные (вторичные) и преовуляторные (третичные) фолликулы. Примордиальные фолликулы размером 50 мкм, заложены ещё до рождения. Они образуются в процессе митотической пролиферациипервичных зародышевых клеток (оогоний), поступивших в зародышевый яичник на 6-й неделе беременности. Они самые многочисленные и мелкие. Их фолликулярные клетки лежат в 1 слой, в центре фолликула – ооцит первого порядка, покоящийся на стадии профазы мейоза. Развитие примордиальных фолликулов ожидает периода полового созревания. К этому времени в яичнике насчитывается около 300 000 фолликулов. Гипофиз начинает вырабатывать фолликулостимулирующий гормон (ФСГ), что стимулирует созревание 5-15 примордиальных фолликулов. Теперь эти фолликулы являются преантральными (первичными) фолликулами (размер — 150-200 мкм). Ооцит начинает расти, внешняя поверхность ооцита покрывается гликопротеинами и гликозаминогликанами, формирующим zona pellucida. Теперь ооцит покрыт уже 2-4 слоями гранулезных клеток, из соединительной ткани формируется оболочка вокруг фолликула. В следующей стадии формируется полость (antrum folliculare), содержащая фолликулярную жидкость (liquor folliculare). Фолликулярные клетки, отвечающие за выработку эстрогена, разделяются на клетки внешней (theca externa) и внутренней (theca interna) оболочки. В это же время эпителиальные клетки фолликула превращаются в гранулёзные клетки, отвечающие за выработку прогестинов. Диаметр антрального (вторичного) фолликула составляет 500 мкм. Во время созревания фолликула клетки внутреннего слоя теки вырабатывают андрогены, которые сквозь базальную мембрану проникают в гранулёзные клетки фолликулярной оболочки и там трансформируются в эстрогены, главным образом, эстрадиол. Таким образом, в фолликулярную полость выделяются эстрогены, а вторичный фолликул становится временным органом эндокринной системы. Образование фолликулярной полости провоцирует быстрый рост фолликула, в течение этого периода диаметр фолликула увеличивается с 1 мм до 16-20 мм непосредственно перед овуляцией. Теперь яйцеклетка расположена на яйценосном бугорке, или яйценосном холмике (cumulus oophorus). Полость составляет большую часть преовуляторного (третичного) фолликула (граафов Примерно за 24 часа до овуляции тека-клетки начинают вырабатывать большое количество эстрогена. Повышенное содержание эстрогена стимулирует выброс лютеинизирующего гормона(ЛГ), который и инициирует овуляцию. В стенке фолликула образуется выпячивание (стигма), которое разрывается, и яйцеклетка выходит из фолликула — происходит овуляция. Если зрелый фолликул не прошёл овуляцию, образуется кистозный фолликул. В норме овуляции подвергается один фолликул, остальные претерпевают инволюцию, образуются атретические тела – блестящая оболочка сморщивается, фолликулярные клетки погибают. После овуляции из фолликула (из гранулёзных и тека-клеток) образуется жёлтое тело, вырабатывающее прогестерон. Прогестерон предотвращает преждевременное отторжение функционального слоя эндометрия (менструация). Если яйцеклетка не была оплодотворена, жёлтое тело прекращает функционировать, уровень прогестерона падает, начинается менструация. Если же произошло оплодотворение, хорион начинает вырабатывать хорионический гонадотропин, который теперь вместо ЛГ стимулирует рост жёлтого тела. Зрелые женские гаметы называют яйцеклетками или яйцами, а отложенные в воду – икрой.
5. Гормональная регуляция оогенеза
В детском возрасте незрелые яичники вырабатывают небольшое количество гормонов, поэтому концентрация эстрогенов в крови низкая. В пубертатный период чувствительность гипоталамо-гипофизарной системы к действию ЛГ и ФСГ снижается. Импульсная секреция гонадотропин-рилизинг-гормона устанавливает суточный ритм секреции ЛГ и ФСГ. В начале каждого менструального цикла секреция ФСГ и ЛГ вызывает развитие первичных фолликулов. У женщин одновременно созревает только один из них (крайне редко — два и больше). Созревающий фолликул в результате совместного действия ЛГ, стимулирующего продукцию андрогенов клетками теки, и ФСГ, стимулирующего ароматизацию андрогенов, секретирует эстрогены (к ним относится эстрадиол), которые по механизму отрицательной обратной связи угнетают секрецию ФСГ. Эстрадиол, вызывающего пролиферацию эндометрия, достигнув достаточно высокой концентрации, по механизму положительной обратной связи усиливает в середине менструального цикла выброс гонадолиберина и повышает чувствительность к нему гипофизарных клеток, секретирующих ФСГ и ЛГ. Концентрация ФСГ в крови остаётся низкой ещё и в результате торможения секреции этого гормона белком ингибином, выделяемым яичниками. ЛГ вызывает овуляцию и лютеинизацию гранулезных клеток фолликула, т.е., повышение секреции ЛГ приводит к овуляции - освобождению яйцеклетки из лопнувшего фолликула. После овуляции клетки гранулёзы превращаются в жёлтое тело, которое, помимо эстрадиола, начинает вырабатывать всё большее количество основного гормона лютеиновой фазы - прогестерона (прогестина). Стероиды, эстрадиол и прогестерон действуют на гипоталамус и гипофиз по механизму отрицательной обратной связи, подавляя во второй половине цикла секрецию ФСГ и ЛГ. Оба гормона действуют также на высшие отделы ЦНС, поэтому у животных половое влечение (либидо) в значительной мере зависит от стадии цикла. Кроме того, существует ряд циклических изменений в других системах организма: ССС, дыхательной, изменения водного баланса, в сенсорных системах и высших отделах ЦНС, что проявляется в динамике сенсорной чувствительности, психоэмоционального напряжения и умственной работоспособности. Если возникает беременность, жёлтое тело продолжает функционировать и секретировать прогестерон, однако на более поздних этапах беременности прогестерон в основном продуцируется плацентой. Если оплодотворение не происходит, высокая концентрация прогестерона в плазме крови по механизму отрицательной обратной связи угнетает активность гипоталамо-гипофизарной системы, тормозится секреция ЛГ и ФСГ, жёлтое тело разрушается, и снижается продукция стероидов яичниками. Наступает менструация, которая длится примерно 5 дней, после чего начинает формироваться новый поверхностный слой эндометрия, и возникает новый цикл. 6. Особенности и отличия сперматогенеза и оогенеза Отличия сперматогенеза от оогенеза у млекопитающих
Кроме этого (!!!), в фазе созревания оогенеза существует 2 блока мейоза: 1. От момента рождения до начала полового созревания – блок профазы 1-го деления мейоза. Снимается благодаря регулирующему действию гормонов гипоталамо-гипофизарно-половой оси. 2. Блок метафазы 2-го деления мейоза – снимается в момент оплодотворения.
7. Происхождение половых клеток В конце XIX в. Вейсман выдвигает теорию о непрерывности зародышевой плазмы. По этой теории существует две плазмы: 1. Соматическая, которая участвует в развитии тела организма; 2. Зародышевая, которая передается непосредственно от половых клеток одного поколения к половым клеткам каждого нового поколения. Теория Нусс-Баума, которая считает, что существует ранняя неравноценность бластомеров. Из одних бластомеров с ограниченными потенциями развиваются соматические клетки, а другие бластомеры – полипотентные – являются источниками половых клеток. Доказательство этой теории привел Бовери в 1939 г., он обнаружил, что уже первые 4 бластомера яйца лошадиной аскариды неравноценны, и один из бластомеров является родоначальником будущих внутренних органов и половых клеток. В 1967 – 1968 гг. Вудлен и Мориц подтверждают это положение и доказывают, что этот процесс ранней дифференцировки половых клеток находится под цитоплазматическим контролем. На свето- и электронно-микроскопическом уровнях доказано, что в одном из бластомеров определяется небольшой участок плотных образований, которые были названы половыми детерминантами, или эктосомами. Предполагается, что эти структуры определяют, или детерминируют, развитие половых клеток в окончательные. По другой теории, источником половых клеток является герментатический эпителий (эпителий половых желез). По следующей теории, помимо половых желез существуют в тех или иных местах латентные половые зачатки.ри некотором условии эти резервные гоноциты оказываются способными развиться в гаметы. Большинство современных исследователей склоняются к мысли, что первичные половые клетки образуются на ранних этапах эмбриогенеза из соматических клеток различных зародышевых листков, но определенных для каждого вида животных.
8. Классификация яйцеклеток по количеству желтка По количеству желтка различают следующие виды яйцеклеток: 1. Полилецитальные — содержат большое количество желтка (членистоногие, рептилии, птицы, рыбы, кроме осетровых). 2. Мезолецитальные — содержат среднее количество желтка (осетровые рыбы, амфибии). 3. Олиголецитальные — содержат мало желтка (моллюски, иглокожие). 4. Алецитальные — не содержат желтка (млекопитающие, некоторые паразитические перепончатокрылые).
9. Классификация яйцеклеток по распределению желтка По распределению желтка в цитоплазме различают следующие виды яйцеклеток:
1. Телолецитальные — желток смещён к вегетативному полюсу яйцеклетки. Противоположный полюс называется анимальным. Сюда относятся некоторые полилецитальные (рыбы, кроме осетровых, рептилии, птицы) и все мезолецитальные яйца (осетровые рыбы, амфибии). 2. Гомо(изо)- лецитальные — желток распределён равномерно. Сюда относятся олиголецитальные ядра (моллюски, иглокожие). 3. Центролецитальные — желток расположен в центре яйцеклетки. Сюда относятся некоторые полилецитальные яйца (членистоногие). Это совершенно особый тип яиц. Вместо анимального и вегетативного полюсов у этих яиц говорят о переднем и заднем полюсах. В центре яйца расположено ядро, а по периферии — ободок свободной от желтка цитоплазмы. Оба этих района — центр и периферия яйца — связаны тонкими цитоплазматическими мостиками, а всё промежуточное пространство заполнено желтком.
|
||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 156; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 13.59.236.219 (0.078 с.) |
 Размножение. Исходные недифференцированные половые клетки – сперматогонии, или гонии делятся путем обычного митоза. Фаза размножения сперматогоний контролируется естественными мутагенами – гормонами аденогипофиза (фоллитропин, интерлекин-1) и гормонами, вырабатываемыми клетками Сертоли и клетками Лейдига. В мужском организме фаза размножения начинается после полового созревания, в репродуктивном возрасте.
Размножение. Исходные недифференцированные половые клетки – сперматогонии, или гонии делятся путем обычного митоза. Фаза размножения сперматогоний контролируется естественными мутагенами – гормонами аденогипофиза (фоллитропин, интерлекин-1) и гормонами, вырабатываемыми клетками Сертоли и клетками Лейдига. В мужском организме фаза размножения начинается после полового созревания, в репродуктивном возрасте.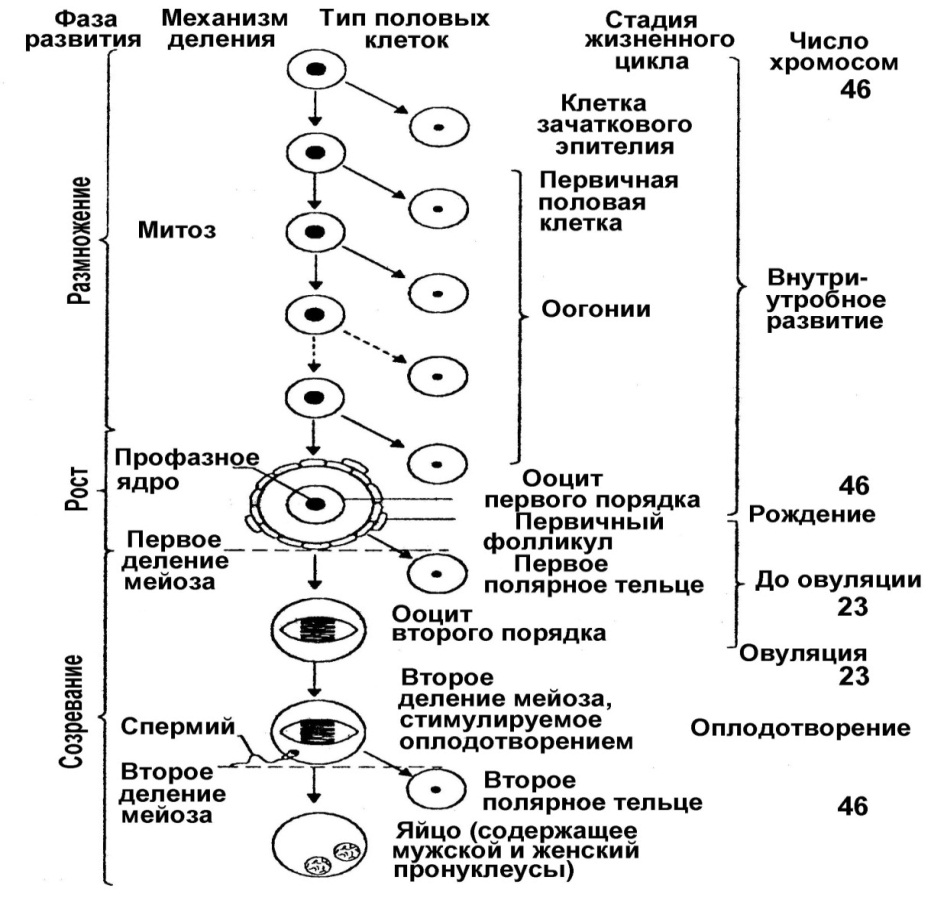 пузырёк), количество фолликулярной жидкости примерно в 100 раз больше, чем в антральном фолликуле.
пузырёк), количество фолликулярной жидкости примерно в 100 раз больше, чем в антральном фолликуле.


